Originally published in the Journal of Mammalogy, 81(3), 666—675.
2000.
By David W Nagorsen and Grant Keddie
Abstract
Although Oreamnos americanus is absent from most Pacific Coast islands, including Vancouver Island, 12,000-year-old skeletal remains were recovered in 2 caves on northern Vancouver Island. The specimens may represent early postglacial immigrants or a relict population derived from a coastal glacial refugium. Limb bones of the fossils are within the size range of modern specimens, suggesting a postglacial origin. O. americanus probably became extinct on Vancouver Island during the early Holocene warming, but inadequacies in the prehistoric faunal record prohibit a determination of a terminal date. The modern distribution of O. americanus on Pacific Coast islands reflects both prehistoric extinctions and low colonization rates across water barriers.
Key words: climatic change, extinction, Holocene, island biogeography, Oreamnos americanus, Pleistocene
Introduction
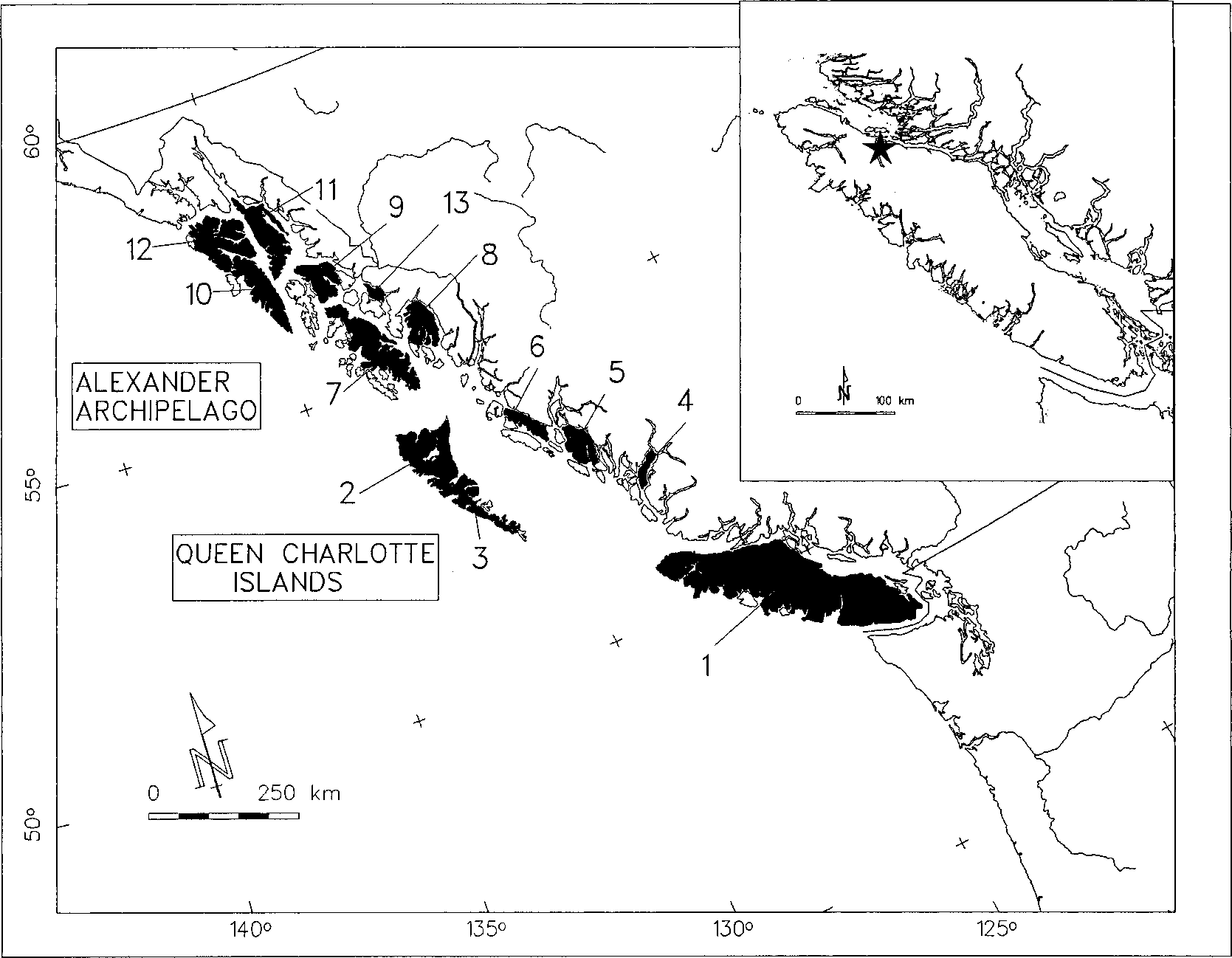
The mountain goat (Oreamnos americanus ) typically inhabits subalpine and alpine habitats associated with cliffs and rock bluffs. In the Pacific Coast region, some populations of O. americanus occupy low- elevation winter ranges near sea level (Hebert and Turnbull 1978). Although widely distributed throughout the coastal mountain ranges of northwestern North America from Washington to southern Alaska, this species is conspicuously absent from the many land-bridge islands of the Pacific Coast (Rideout and Hoffmann 1975). Pitt Island on the central coast of British Columbia (Fig. 1) supports the only natural, extant insular population (Gosnell 1911; McCabe and Cowan 1945). A solitary mountain goat was sighted in 1995 on nearby Princess Royal Island. There are no established natural populations of O. americanus on any of the Alaskan islands, although mountain goats were introduced successfully to Ba- ranof, Revillagigedo, and Kodiak islands, and an animal was observed on Wrangell Island for several years (Klein 1965; MacDonald and Cook 1996). Records from Cul- ross and Bainbridge (Klein 1953, 1965), 2 small nearshore islands (,500 m from mainland) in Prince William Sound, probably represent transients, as these islands are too small to support sustainable populations. Absence of mountain goats on Vancouver Island is curious. With an area of 33,000 km2, rugged mountains some .2,100 m elevation, and nearly 125 km2 of alpine area (British Columbia Ministry of Environment, Lands and Parks, in litt.), Vancouver Island clearly has potential habitat for O. americanus. Size of the alpine zone is a conservative estimate of habitat for O. americanus because coastal populations of mountain goats also exploit lower elevations for their critical winter range (Hebert and Turnbull 1977).
Klein (1965) attributed the distributional pattern of O. americanus on the Alaskan islands to colonization effects and the species’ poor dispersal abilities across water barriers. Nonetheless, Quaternary fossils found on Prince of Wales Island (Heaton et al. 1996) demonstrate that postglacial extinctions also have occurred on islands of the Alexander Archipelago. The relative contributions of colonization and extinction to the modern insular distribution of O. americanus cannot be determined solely from Recent distributional patterns, and it is essential to examine the species’ prehistoric range. Unfortunately, with an inadequate fossil record and poor representation in archaeological sites, the historical biogeography of O. americanus in northwestern North America is largely speculative (Lyman 1988). Quaternary fossils are known from only 2 localities in the Sierra Nevada and 5 sites in the continental interior (Harington 1971; Mead and Lawler 1994). The species also is poorly represented in archaeological sites, and any interpretation of archaeofaunal remains is hindered by sampling biases (Lyman 1995).
Caves in karst landforms have yielded important Quaternary mammalian remains on islands of the Alexander Archipelago (Heaton et al. 1996) and Vancouver Island (Nagorsen et al. 1995). During a preliminary faunal analysis of caves on northern Vancouver Island, cavers from the Vancouver Island Cave Exploration Group recovered postcranial material from 2 individuals of O. americanus. They represent the 1st Pleistocene records of this species for the Pacific Coast and provide the 1st evidence for extinction of a Recent mammal from Vancouver Island. Herein, we describe the specimens, compare their size with modern specimens, and discuss the biogeographic significance.
Materials and Methods
Remains of mountain goats were found in Resonance and Pellucidar, 2 caves east of Nimp- kish Lake on northern Vancouver Island, British Columbia (Fig. 1). The caves are about 5 km apart and are situated at 750-810 m above mean sea level in the Coastal Western Hemlock biogeographic zone (Meidinger and Pojar 1991). Vegetation is forested mostly with western hemlock (Tsuga heterophylla) and some western red cedar (Thuja plicata). Caves are part of an extensive karst landscape (Quatsino Limestone formation) on the lower slopes of the Hankin Range. Relief is gently rolling. The highest mountain of the range (Mount Hoy) has open subalpine habitat but lacks vertical cliffs or shear rock faces that are characteristic of O. americanus escape terrain. The nearest mountains with typical habitat for mountain goats are the Bonanza Range, 30 km SE, and the Karmutzen Range, 10 km W, of our study area.
Resonance Cave (810 m elevation) consists of a narrow 100-m-long horizontal tunnel that ends at a 15-m vertical drop to a small chamber. Seven bones of O. americanus were located in the main tunnel about 30 m from the entrance in a shallow (10 cm) pool of silty deposits on the cave floor. No other vertebrates were found in association with the specimens of mountain goats. We found no archaeological evidence (cultural material or fire pits) of human occupation. Nevertheless, because the cave is readily accessible and a popular recreation site, archaeological evidence could have been removed.
Pellucidar Cave (750 m elevation) is a large (total surveyed length 1,400 m) complex cave system associated with an underground stream. A 12-m submerged chamber (sump) divides the main passageway into 2 sections (Pellucidar I and Pellucidar II). A secondary passage in Pellucidar II about 140-m long leads to a 20-m- wide chamber with extensive rock rubble that appears to be a collapsed former entrance. Access to this chamber was through a small opening to the surface excavated recently by cavers. A femur of a mountain goat was found in the chamber among the surface rock rubble. A preliminary survey of surface bones revealed mule deer (Odocoileus hemionus), black bear (Ursus americanus), marmot (cf. Marmota vancouver- ensis), Peromyscus, Microtus, Sorex, Myotis, red-tailed hawk (Buteo jamiacensis), and a large amphibian (cf. Bufo) in Pellucidar II. Large deposits (2 m high) of fine-grained sediments in Pellucidar II that are associated with the stream have not been excavated and analyzed for microvertebrate remains. Bones of M. vancouver- ensis and Peromyscus also were found in calcareous mud on the floor about 50 m from the entrance of Pellucidar I. We found no archaeological evidence for human occupation of Pellucidar Cave.
We extracted small plugs of bone (1-2 g) from the 2 femurs with a diamond core-bit drill for radiocarbon dating. Material was dated by the Isotrace Radiocarbon Laboratory at the University of Toronto from collagen using the accelerator mass spectrometry method.
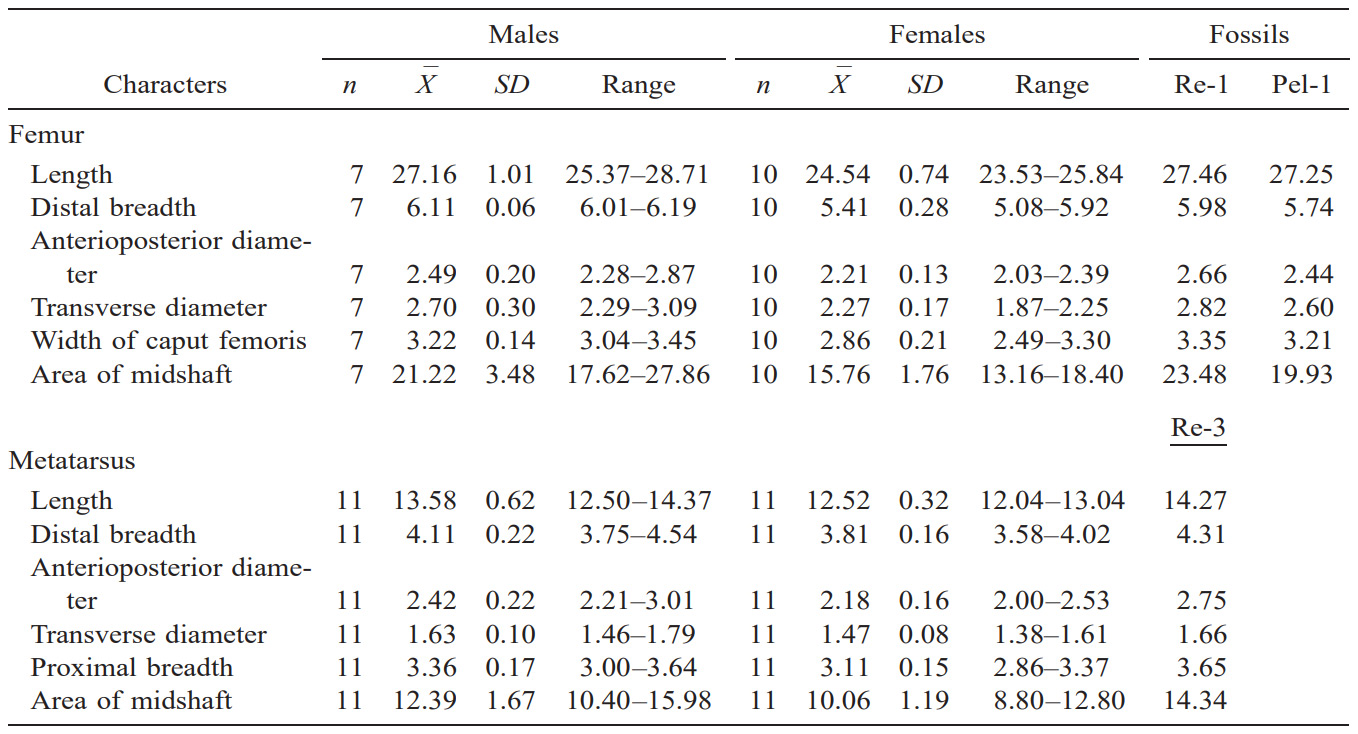
Size analyses were based on femurs and metatarsals from the 3 fossil specimens and 22 modern O. americanus. The few available skeletal specimens prohibited a rigorous analysis of geographic variation in modern O. americanus, but collecting localities span the entire geographic range (Appendix I). Only adults were used (complete epiphyseal fusion). Because O. americanus demonstrates sexual dimorphism in body (Rideout and Hoffmann 1975) and cranial (Cowan and McCrory 1970) measurements, we restricted our analyses to specimens of known sex.
We took 5 femoral (right side) and 5 metatarsal (left side) measurements to the nearest 0.01 cm using Mitutoyo dial calipers: length of femur, greatest length of femur from caput femor- is; distal breadth, greatest breadth of distal end of femur; anterioposterior diameter, anterioposterior diameter of femur measured at midshaft; transverse diameter, transverse diameter of femur measured at midshaft; width of head, greatest width of caput femoris; length of metatarsus, greatest length of metatarsus; distal breadth, greatest breadth of metatarsus at distal end; anterioposterior diameter, anterioposterior diameter of metatarsus measured at midshaft; transverse diameter, transverse diameter of metatarsus measured at midshaft; and proximal breadth, greatest breadth of metatarsus at proximal end. Crosssectional areas of femurs and metatarsals at midshaft were derived from the area of an ellipse using anterioposterior and transverse diameters as the major and minor axes. Such measurements previously were illustrated and defined by Scott (1985) and von den Driesch (1976). We compared individual measurements of fossils with means of modern samples using a 2-tailed Student’s t-test.

We used 13 major islands from the Alexander Archipelago and British Columbia coast (Fig. 1) to assess effects of isolation (distance to mainland) and area on the insular distribution of O. americanus: Admiralty, Baranof, Chichagof, Graham, King, Kupreanof, Moresby, Pitt, Princess Royal, Prince of Wales, Revillagigedo, Vancouver, and Wrangell. They range from 540 to 33,000 km2, have high elevational relief (970-2,200 m), and demonstrate a wide range of isolation (0.5-142.0 km from the mainland).
Results
The 7 specimens in Resonance Cave included a complete right femur (Fig. 2), complete left metatarsus, the proximal section of a left rib, an ossified cartilaginous rib fragment, 3 incomplete vertebrae (2 thoracic, 1 lumbar), left astragalus, and sesamoid bone. Calcareous encrustrations were evident on the femur. Given their proximity and similar stage of maturation, the 7 specimens probably represented a single animal. The fused epiphyses of the femur and metatarsal with their robust shafts and pronounced wear on their articular surfaces suggested an old individual. The single specimen of O. americanus from Pellucidar Cave was a right femur with most of the greater trochanter missing (Fig. 2). The fused epiphyses suggested a mature animal. We observed no carnivore tooth punctures, rodent gnaw marks, or evidence of human modification on bones.
Both femurs were radiocarbon dated as late Pleistocene. The Resonance Cave sample (RBCM RE-1, lab no. TO-6072) was 12,200 6 190 radiocarbon years old; the Pellucidar Cave sample (RBCM PELII-1, lab no. TO-5006) was 12,070 6 70 radiocarbon years old.
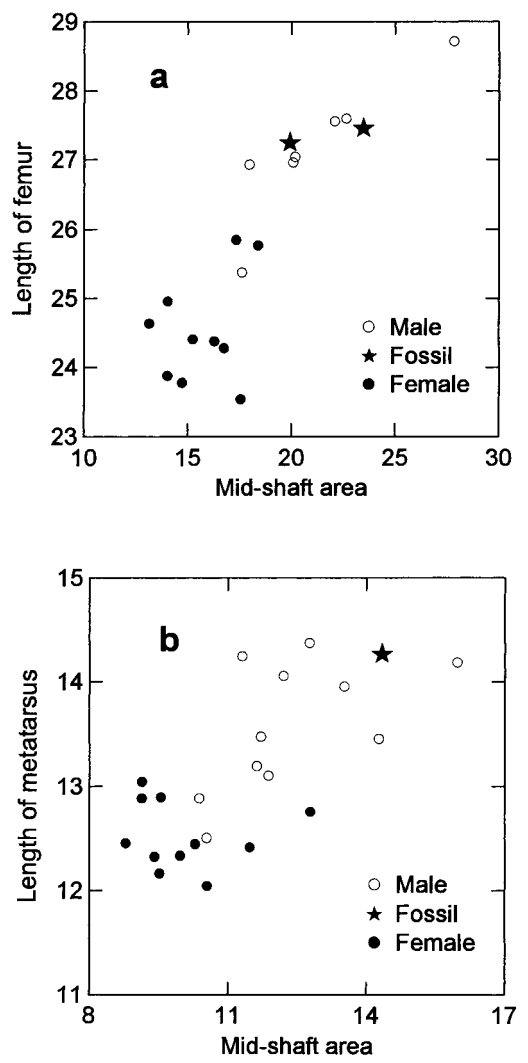
Modern O. americanus demonstrate pronounced sexual dimorphism in size of the femur and metatarsus (Table 1) with means of males larger (P , 0.01) for all 12 measurements of limb bones. The fossil limb bones are long and robust (Figs. 3a and 3b). The 6 femoral measurements of the 2 fossils exceed the maximum values for modern females, but none differ from means of modern males (P . 0.05). The fossil metatarsal is also large, exceeding maximum measurements for any modern female and approaching the maximum size of modern males, although none of the 6 measurements differ from means of modern males (P . 0.05).
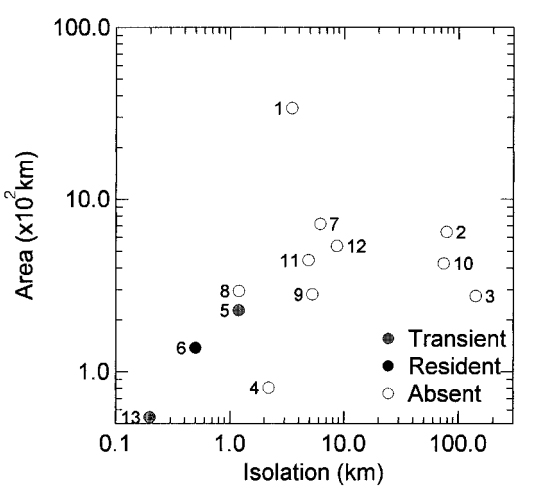
The modern distribution of O. americanus among 13 large islands in southeastern Alaska and British Columbia strongly reflects isolation effects (Fig. 4) with mountain goats only occupying islands ,1.5 km from the nearest mainland. Pitt Island, which has the only known resident population, is separated from the mainland by a narrow channel of about 0.5 km. Similarly, sightings of solitary animals that probably represent transients are restricted to nearshore islands: Wrangell (0.2 km) and Princess Royal (1.2 km). A noteworthy pattern is that the largest islands in the coastal archipelagos of southeastern Alaska and British Columbia are the most isolated islands with some >70 km from the nearest mainland. Vancouver Island is an outlier in size with an area >3 times that of other large islands.
Discussion
The 12,000-year-old radiocarbon dates for the 2 O. americanus from Vancouver Island correspond to the early postglacial. Our study area was covered with advancing ice of the Fraser glaciation 20,000 years ago with ice reaching .1,300 m thick in some areas (Clague 1981; Howes 1981). Maximum glaciation on northern Vancouver Island occurred about 15,000 years ago, but that period was brief. Deglaciation began by 13,000 years ago at Port McNeill (Howes 1983), and vegetation appeared as early as 13,600 years ago at Bear Cove (Hebda 1983), 2 sites on the northeast coast of Vancouver Island. By 12,000 years ago, the 2 cave sites were presumably ice free, although glaciers may have persisted in the Nimpkish River Valley (Howes 1981). The climate during this early postglacial period was cool and dry with plant communities dominated by lodgepole pine (Pinus con- torta—Hebda 1983, 1995). Lowland landscapes probably were a pine parkland with open meadows. Presence of marmots in Pellucidar Cave is consistent with an open parkland habitat. This early postglacial habitat resembled the winter range of modern O. americanus living in coastal areas (Hebert and Turnbull 1978). However, the lack of precipitous cliffs in the immediate vicinity of the caves is curious given the importance of this type of habitat for O. americanus in providing escape terrain from predators. Possibly, potential predators of mountain goats, such as the gray wolf (Ca- nis lupus) and mountain lion (Puma con- color), were absent from Vancouver Island in this early postglacial period.
The O. americanus that inhabited Vancouver Island in the late Pleistocene may have been a relict population that survived the Fraser glaciation in refugia associated with nunataks or large areas of the continental shelf that were exposed during the late Pleistocene (Josenhans et al. 1995).
Heaton et al. (1996) and Byun et al. (1997) argued that some mammals persisted in coastal refugia throughout the last glaciation. Alternatively, our fossils could represent a population derived from postglacial immigrants that followed the receding Cor- dilleran ice sheet on the mainland and colonized the island by ice bridges or exposed sea floor that briefly linked Vancouver Island to the mainland in the early postglacial (Clague 1981). With its efficient insulating pelage and ability to traverse small glaciers or ice fields (Klein 1953), O. americanus would be well adapted to survive in a periglacial environment. This postglacial colonization scenario would be consistent with the dispersal model for O. americanus proposed by Lyman (1988) for western Washington.
Lack of fossils of O. americanus from the Pacific Coast of North America (Mead and Lawler 1994) prohibits a rigorous evaluation of these competing hypotheses. Pronounced morphological differentiation in body size, such as dwarfism (Lomolino 1985), would be consistent with long-term isolation and a possible refugial origin. Changes in body size can evolve rapidly in insular mammalian populations. For example, in a population of red deer (Cervus ela- phus) isolated on Jersey Island in the English Channel, diameters of limb bones decreased by about 56% and estimated body weight by .80% within a period of only 6,000 years (Lister 1989). Because most museum specimens lack data for body weight, we could not assess the relationship of body mass and size of limb bones in our sample of modern O. americanus. Scott (1985) demonstrated that lengths and midshaft areas of femurs and metatarsals scaled closely with body mass in 108 species of Bovidae, and we assume that size and robustness of limb bones are indicators of body size of mountain goats. Limb bones of the Pleistocene O. americanus from Vancouver Island show no evidence of dwarfism but tend to be large, although they do not exceed the size of large modern males. No contemporary fossils from the coastal mainland exist for comparison, but it appears that the O. americanus that inhabited the continental interior in the late Pleistocene were similar in size to modern animals. Three metacarpals and a single metatarsal of Rancholabrean age from Booth Canyon, Idaho, used in the analyses by Mead and Lawler (1994) were similar in size to modern animals. Lack of strong morphometric differentiation from mainland forms is concordant with a postglacial origin for the population on Vancouver Island.
Why O. americanus became extinct on the largest island in the eastern North Pacific is not clear Mead and Lawler (1994) speculated that a shift to hot summers during the late glacial-early postglacial transition caused extinction of Harrington’s mountain goat (Oreamnos harringtoni) in the southwestern United States about 11,000 years ago. Rapid climatic and associated vegetational changes in the Holocene coupled with vulnerable life-history traits of coastal mountain goats, such as low recruitment rates and small size of groups (Hebert and Turnbull 1977), would contribute to stochastic extinctions of insular populations of O. americanus. The most pronounced climatic change after glaciation on Vancouver Island was a rapid warming trend about 10,000-7,500 years ago with mean annual temperatures 2-48C warmer than today (Hebda 1995). With warmer temperatures and reduced precipitation in winter, the tree-line shifted in the early Holocene, advancing 60-130 m higher than modern elevations (Rochefort et al. 1994) reducing open-meadow habitat, and increasing habitat fragmentation for montane species such as O. americanus.
Our hypothesis of an early Holocene extinction, however, is speculative because the terminal date for O. americanus on Vancouver Island is unknown. Reports from early explorers (Schultz 1994) and the 1st zoological surveys (Swarth 1912) strongly indicate that mountain goats were absent from Vancouver Island in historical time. In 1924, an unsuccessful attempt was made to introduce the species to the island (Lloyd 1925). Aboriginal groups on Vancouver Island used mountain goat horns and wool, but ethnographic data suggest that these items were obtained by trading with mainland peoples (Schultz 1994). Faunal remains of O. americanus are lacking for the entire Holocene of Vancouver Island, but this may simply reflect sampling and the inadequate prehistoric faunal record. The mammalian paleontological record from Vancouver Island consists of late Pleistocene fossils of extinct species, such as the helmeted musk ox (Symbos cavifrons), imperial mammoth (Mammuthus imperator), mastodon (Mammut americanum), and horse (Equus—Harington 1975). Postglacial paleontological remains are limited to several opportunistic finds of bison (cf. Bison bison occidentalis) in bogs (Harington 1996) and cave deposits reported herein and by Nagorsen et al. (1995). Holocene mammalian remains are common in archaeological sites, but the archaeological record for Vancouver Island has major geographic and temporal biases with most archaeofaunal remains associated with lowland coastal occupation sites that date from the past few thousand years (Mitchell 1990). Lyman (1995) demonstrated that archaeofaunal remains of O. americanus in Washington and Oregon were in montane cave and rock shelter sites in proximity to habitat of mountain goats. Only 4 subalpine rockshelters with faunal remains are known from the island; all are of late Holocene age (Nagorsen and Keddie 1996). With the current gaps in the prehistoric faunal record, it is conceivable that O. americanus persisted well into the Holocene on Vancouver Island.
Whatever the timing of the disappearance of O. americanus from Vancouver Island, its failure to persist on an island of 33,000 km2 suggests that it is a poor survivor on islands. A large body size, requirements for alpine habitat, and fragmented distributional patterns are life-history traits that would make this species prone to extinction on islands. Although there are anecdotal observations of O. americanus swimming across small lakes and swift glacial streams (Klein 1953), there are no empirical data on this species’ ability to swim in seawater. Nevertheless, its current insular distribution is correlated with isolation effects. We interpret the modern insular distribution of O. americanus as the result of both prehistoric extinctions and low dispersal abilities across water barriers. Following a rapid rise in sea levels in the early postglacial (Clague 1981), relictual populations of O. americanus derived either from refugia or early postglacial colonization from the mainland may have been isolated on a number of the large islands of southeastern Alaska and British Columbia. Evidently, all went extinct except for Pitt Island. Known since the early 1900s (Gosnell 1911), this population is separated from the mainland by a narrow channel of only 500 m and probably is maintained by recurrent immigration from the mainland. On the central coast of British Columbia, some populations of O. americanus inhabit winter ranges near sea level (Hebert and Turnbull 1978), thus providing a pool of potential colonizers for nearshore islands. Most other large islands of British Columbia and Alaska with potential habitat for O. americanus are simply too isolated for over-water immigration. The northeastern coast of Vancouver Island, for example, is connected to the mainland by a series of stepping-stone islands in Johnstone Strait, but they all have low relief and lack suitable montane habitat. Shortest distance to the mainland in this region is about 3.5 km, and the nearest known population of O. americanus is in the Knight Inlet area about 35 km from Vancouver Island. Other large islands, such as Graham and Moresby in the Queen Charlotte Islands (Haida Gwaii) Archipelago and Admiralty, Baranof, Chichagof, and Prince of Wales in the Alexander Archipelago, are even more isolated (5-140 km) from the mainland.
Postglacial colonization (Klein 1965) undoubtedly played a major role in determining the mammals that currently occupy the land-bridge islands of coastal Alaska and British Columbia. The late Pleistocene remains of O. americanus from Vancouver Island, the discovery of Holocene fossils of 4 extant mammalian species now absent from Prince of Wales Island (Heaton et al. 1996), and the disappearance of caribou (Rangifer tarandus) on the Queen Charlotte Islands in the early 1900s (Banfield 1962) demonstrate that postglacial extinctions also shaped these faunas. Presumably, the early postglacial on these islands was a period of high turnover of species resulting from pronounced climatic and geologic changes and competition effects. The colonization history and precise timing of extinctions is poorly known, and more dated fossil remains are essential to unravel the complex biogeographic history of coastal British Columbia and Alaska.
Acknowledgments
We are indebted to P. Curtis, B. Nasby, and S. McNay, who discovered the fossils, guided us through the caves, and assisted with collecting specimens. D. Klein provided a copy of his M.S. thesis, D. Fowle measured specimens, S. Crockford and B. Wigen verified our identifications, K. Brunt provided data on habitat and distributions of mountain goats in British Columbia, and D. Shackelton commented on a draft. We thank curators and collection staff at the following institutions for allowing us to measure specimens in their care or providing loans: American Museum of Natural History (AMNH), New York; Cowan Vertebrate Museum University of British Columbia (UBC), Vancouver; Canadian Museum of Nature (CMN), Ottawa; Museum of Vertebrate Zoology (MVZ), University of California, Berkeley; Provincial Museum of Alberta (PMA), Edmonton; James R. Slater Museum of Natural History, University of Puget Sound (UPS), Tacoma; United States National Museum of Natural History (USNM), Washington, D.C.; and University of Alaska Museum (UAM), Fairbanks. Research was funded by the Royal British Columbia Museum (RBCM) and Friends of the Royal British Columbia Museum.
Literature Cited
Banfield, A. W. F 1962. The disappearance of the Queen Charlotte Islands’ caribou. Bulletin of the National Museum of Canada 185:40-49.
Byun, A. S., B. F Koop, and T. E. Reimchen. 1997. North American black bear mtDNA phylogeogra- phy: implications for morphology and the Haida Gwaii refugium controversy. Evolution 51:16471653.
Clague, J. J. 1981. Late Quaternary geology and geochronology British Columbia. Part 2. Summary and discussion of radiocarbon-dated Quaternary history. Geological Survey of Canada, Paper 80-35:1-41.
Cowan, I. M., and W. McCrory. 1970. Variation in the mountain goat, Oreamnos americanus (Blain- ville). Journal of Mammalogy 51:60-73.
Gosnell, R. E. 1911. The year book of British Columbia and manual of provincial information. Government of British Columbia, Victoria, British Columbia, Canada.
Harington, C. R. 1971. A Pleistocene mountain goat from British Columbia and comments on the dispersal history of Oreamnos. Canadian Journal of Earth Sciences 8:1081-1093.
Harington, C. R. 1975. Pleistocene muskoxen (Sym- bos) from Alberta and British Columbia. Canadian Journal of Earth Sciences 12:903-919.
Harington, C. R. 1996. Quaternary animals: vertebrates of the ice age. Pp. 259-273 in Life in stone (R. Ludvigsen, ed.). University of British Columbia Press, Vancouver, British Columbia, Canada.
Heaton, T H., S. L. Talbot, and G. F Shields. 1996. An ice refugium for large mammals in the Alexander Archipelago, southeastern Alaska. Quaternary Research 46:186-192.
Hebda, R. J. 1983. Late-glacial and postglacial vegetation history at Bear Cove bog, Northeast Vancouver Island, British Columbia. Canadian Journal of Botany 61:3172-3192.
Hebda, R. J. 1995. British Columbia vegetation and climate history with focus on 6 KA BP. Geographie physique et Quaternaire 49:55-79.
Hebert, D. M., and W. G. Turnbull. 1978. A description of southern interior and coastal mountain goat ecotypes in British Columbia. Pp. 126-146 in Proceedings of the first international mountain goat symposium (W. Samuel and W. G. Macgregor, eds.). Province of British Columbia, Ministry of Recreation and Conservation, Fish and Wildlife Branch, Victoria, British Columbia, Canada.
Howes, D. E. 1981. Late Quaternary sediments and geomorphic history of north central Vancouver Island. Canadian Journal of Earth Sciences 18:1-12.
Howes, D. E. 1983. Late Quaternary sediments and geomorphic history of northern Vancouver Island, British Columbia. Canadian Journal of Earth Sciences 20:57-65.
Josenhans, H. W., D. W. Fedie, K. W. Conway, and J. V. Barrie. 1995. Postglacial sea levels on the western Canadian continental shelf: evidence for rapid change, extensive subaerial exposure, and early human habitation. Marine Geology 125:73-94.
Klein, D. R. 1953. A reconnaissance study of the mountain goat in Alaska. M.S. thesis, University of Alaska, Fairbanks.
Klein, D. R. 1965. Postglacial distribution patterns of mammals in the southern coastal regions of Alaska. Arctic 18:7-20.
Lister, A. M. 1989. Rapid dwarfing of red deer on Jersey in the last interglacial. Nature 342:539-542.
Lloyd, H. 1925. The introduction of the muskrat and the Rocky Mountain goat on Vancouver Island. The Canadian Field-Naturalist 39:151-152.
Lomolino, M. V. 1985. Body size of mammals on islands: the island rule reexamined. The American Naturalist 125:310-316.
Lyman, R. L. 1988. Significance for wildlife management of the late Quaternary biogeography of mountain goats (Oreamnos americanus) in the Pacific Northwest, USA. Arctic and Alpine Research 20: 13-23.
Lyman, R. L. 1995. Determining when rare (zoo)-ar- chaeological phenomenon are truly absent. Journal of Archaeological Method and Theory 2:369-424.
MacDonald, S. O., and J. A. Cook. 1996. The land mammal fauna of Southeast Alaska. The Canadian Field-Naturalist 110:571-598.
McCabe, T T., AND I. M. Coaaaa.i 1945. Peromyscus maniculatus macrorhinus and the problem of insularity. Transactions of the Royal Canadian Institute 25:117-216.
Mead, J. I., and M. C. Lawler. 1994. Skull, mandible, and metapodials of the extinct Harrington’s mountain goat (Oreamnos harringtoni). Journal of Vertebrate Paleontology 14:562-576.
Meidinger, D., aad j. Poiar (eds.). 1991. Ecosystems of British Columbia. British Columbia Ministry of Forests, Special Report Series 6:1-330.
Mitchell, D. 1990. Prehistory of the coasts of southern British Columbia and northern Washington. Pp. 340-358 in Handbook of North American Indians: Northwest coast (W. Suttles, ed.). Smithsonian Institution Press, Washington, D.C., 7:1-777.
Nagorsen, D. W., aad g. Keddie. 1996. Vancouver Island marmot bones from subalpine caves: archaeological and biological significance. British Columbia Ministry of Environment, Lands and Parks, British Columbia Parks Occasional Paper 4:1-56.
Nagorsen, D. W., G. Keddie, and R. J. Hebda. 1995. Early Holocene black bears, Ursus americanus,from Vancouver Island. The Canadian Field-Naturalist 109:11-18.
Rideout, C. B., and R. S. Hoffmann. 1975. Oreamnos americanus. Mammalian Species 63:1-6.
Rochefort, R. M., R. L. Little, A. Woodward, and D. L. Peterson. 1994. Changes in sub-alpine tree distribution in western North America: a review of climatic and other casual factors. Holocene 4:89100.
Schultz, S. 1994. Review of the historical evidence relating to mountain goats in the Olympic Mountains before 1925. Pp. 256-293 in Mountain goats in Olympic National Park: biology and management of an introduced species (D. B. Houston, E. G. Schreiber, and B. B. Moorhead, eds.). United States Department of the Interior, National Park Service, Scientific Monograph NPS/NOLYM/NRSM-94/25: 1-295.
Scott, K. M. 1985. Allometric trends and locomotor adaptations in the Bovidae. Bulletin of the American Museum of Natural History 179:197-288.
Swarth, H. S. 1912. Report on a collection of birds and mammals from Vancouver Island. University of California Publications in Zoology 10:1-124.
VON DEN Driesch, A. 1976. A guide to the measurement of animal bones from archaeological sites. Peabody Museum of Archaeology and Ethnology, Peabody Museum Bulletin 1:1-137.
Submitted 17 March 1999. Accepted 19 November 1999.
Associate Editor was Troy L. Best.
Appendix I
Specimens examined.—Catalog numbers and museum acronyms (see Acknowledgments) for specimens measured. Femurs of modern O. americanus (17). Alaska: no locality (128105 AMNH), Cook Inlet (4344 MVZ), Saint Matthews Bay (19764 UAM), Tracy Arm (130223 AMNH). Alberta: Alberta Game Farm (Z73.26.1 PMA), Banff National Park (774 UBC, 3826 UBC), Eagle Nest Pass (122670 AMNH), Grand Cache (Z72.89.1 PMA), Jasper National Park (41054 CMN, 75079 CMN). British Columbia: Coldfish Lake (4546 UBC), Ingenika River (10207 RBCM), Isaacs Lake (43909 MVZ). Montana: Bridge Pass (2592 USNM), National Bison Range (24691 UAM). Washington: Mount Rainier (24161 UPS).
Femurs of fossil O. americanus (2). British Columbia: Resonance Cave, Vancouver Island (RE-1 RBCM), Pellucidar Cave, Vancouver Island (Pel-1 RBCM).
Metatarsals of modern O. americanus (22). Alaska: no locality (128105 AMNH), Cook Inlet (4344 MVZ), Saint Matthews Bay (19764 UAM), Tracy Arm (130223 AMNH). Alberta: Banff National Park (774 UBC, 775 UBC, 3826 UBC, 238665 USNM, 240998), Eagle Nest Pass (122670 AMNH), Grand Cache (Z72.64.1 PMA, Z72.89.1 PMA), Jasper National Park (41054 CMN, 75079 CMN), Wilmore Wilderness Park (Z73.7.1 PMA). British Columbia: Coldfish Lake (4546 UBC), Ingenika River (10207 RBCM), Isaacs Lake (43909 MVZ), Sitcum Creek (8956 UBC). Montana: National Bison Range (24690 UAM, 24691 UAM). Washington: Mount Rainier (24161 UPS).
Metatarsals of fossil O. americanus (1). British Columbia: Resonance Cave, Vancouver Island (RE-3 RBCM).