By Grant Keddie. 1995.
Primarily Vancouver Island Marmot with Bear and Deer.
Preface
This document is intended as a working technical manuscript from which information will be extracted for other reports and publications related to the Vancouver Island Marmot Cave Project undertaken jointly by Mammologist Dave Nagorsen and myself. The bone element database, metric summaries, analysis and biogeographical data on modern marmots is not included here.

Introduction
This study represents the physical examination of thousands of discarded bone remains found on the surface in caves or rock shelters at four high elevation mountain hunting locations: the Mariner Cave (elevation 1220M), on Mariner Mountain and the Golden Hinde Rock Shelter (1420m) on Golden Hinde Mountain, both in Strathcona Provincial Park; Clayoquot Cave (1220M) on the Clayoquot plateau; and Limestone Rock Shelter (1205M) on Limestone Mountain south of Alberni Inlet.
The Clayoquot and Golden Hinde remains appear to each represent one episode of marmot hunting. The remains from the other localities represent multiple episodes of hunting mostly marmot, but also bear deer and a few marten, squirrel and blue grouse.
The presence of marmot bones in these sites allows us to examine the biogeography of marmots with a greater time perspective. The biological identification and analysis of the bones can lead to determining the number of animals that were hunted and their age distribution. From this we can determine the probable season of hunting and the possible impact of human predation.
The study of these bone assemblages presents a unique opportunity to reconstruct human events at limited time periods. The number, completeness of the bones, and the limited spatial context in which the bone assemblages occur allows for greater certainty in making suppositions about the sequence of events occurring at these locations.
This is in contrast to the more fragmentary nature of bones found in coastal shellmiddens containing bones of numerous species and complex soil features over large areas. The context of the latter bones is difficult to interpret due to in existence of so many variables.
Objectives
The main objective of this examination of the marmot, deer and bear bones is to reconstruct the finer details of human-animal interactions in these specialized hunting sites. This documentation will provide the basis for suppositions on how the animals were processed and what parts were removed or transported to other locations. The activities of this report include:
(1) Documenting and interpreting the visible evidence of human induced bone modifications resulting from skinning, dismembering, filleting, or other activities involving the intentional alteration of bone such as marrow extraction or fracturing of bone to obtain raw materials for tool manufacture. This includes observing those bones and parts that are not modified.
(2) Making observations on the types, parts, and proportions of bone elements that are both present and absent.
(3) Making observations on the distribution of bone elements within the site locations and speculating on why the bones are distributed the way they are within the sites.
(4) Observing the modification of bones those results from natural nonanimal agents. In order to fully understand the nature of the human alterations to the bones it is necessary to also observe changes that result from geochemical modification, root action, weathering, rock fall abrasion or changes that are a result of the weakness of the biological structure of the bone affected by these natural phenomena.
(5) Observing the modification of bones those results from non-human animal activities.
(6) Determining what tools may have been used in the processing of the animals.
Bone Examination And Analysis
All bones (with the exception of those from Golden Hinde Rockshelter) were examined by the naked eye as well as under a microscope at both 10 and 25 power. During the initial process of examination all bones were viewed twice. Later many bones were examined numerous times to recheck for the presence or absence of certain modifications. The Golden Hinde rockshelter bones were examined in the field with a 10x hand lens.
Some bones were reconstructed by identifying fragmentary parts of the same bone. These were sometimes identified after bone numbering – resulting in several numbers representing parts of the same bone.
In identifying the minimum number of individuals, especially in the case of cranium and mandibles, an extra effort was made to identify fragments that were defined as less than half of an element on the electronic database, but are mutually exclusive of all similar elements based on a close physical comparison. This generally resulted in a slightly higher MNI count for some bones, and especially for craniums in the Limestone cave site.
No attempt is made here to systematically document the large number of non-human induced marks or features on the bones. These other features, that may be misinterpreted as cut marks by the casual observer, were examined to assure their separation from genuine cut marks.
In order to determine if the set of bones retrieved is representative of the total number of animals killed an attempt was made to determine the minimum number of each animal represented by the largest number of a single discrete bone (e.g. left mandibles) for each site. From this number the total number of each bone element that should be present to account for all of the processed animals was determined. This allows us to observe what bone elements are missing from each site.
Non Animal/Human Modifications
Natural Bone Features
There are many natural features of bones that can be mistaken as cut marks. These include smooth grooves from periosteal blood vessels and bone laminae exposed by abrasive effects. Natural grooved areas, which vary considerably from specimen to specimen, often are associated with small oscicles at one end.
Geochemical Modification
In most archaeological sites it would be expected that chemical erosion and other natural processes would result in the disappearance of most of the least dense bone elements. These would be under-represented relative to the densest elements. In the sites examined here this is not generally the case, but there are different levels of exposure present in the site locations and this is reflected in different levels of preservation.
The more delicate skull parts have been affected by natural events, and partial erosion of some elements such as the thin edges of scapula, has occurred. Erosion of bone elements was more severe in those bones that were in direct contact with soil or on the bottom of bone piles as opposed to those lying on rock surfaces or on top of bone piles. The more fragmentary skulls from the Limestone rock shelter were mostly in the litter mat and more exposed to the elements than those in other sites. In Mariner cave delicate inter-nasal bones of deer laying on rock surfaces were well preserved. Deer scapula and innominate bones show breakages mostly along the edges due to chemical erosion and/or crushing.
Some bones in Mariner cave were under rock falls and are likely those that show a lower degree of chemical erosion and a lighter colouration on the scrape surfaces. Some of these rock abrasion scrapes are characterized by gouged scares at one end.
Carnivore, Bird And Rodent Modifications
Bones can be altered by animal trampling, carnivore gnawing and rodent chewing activity and animal removal of the bones.
The finding of modern grouse dung and feathers in the small alcove containing the bones at the Limestone Rockshelter suggests the possibility of animal assistance in the scattering of bones. The fragmentation of the cranium at this site may have been assisted by bird trampling and scratching.
Rodents chew bones for the calcium content and to keep their teeth filed down. Many of the bones from all of the sites have Rodent canine chew marks. These are often represented by two parallel grooves with rounded to flattened bottoms. Sometimes these grooves are furrowed or scooped out in the softer bone tissue. Chew marks can include a multiple series of adjoining cuts or grooves. The rodent chew marks represent a wide time period. Some are old weathered chew marks and others, showing fresh cuts with no weathering, are obviously of a recent nature. Many bones had both rodent chew marks and knife butcher marks.
Few obvious carnivore chew marks were observed. Those observed sometimes had a straight cut mark, like a knife cut, but were structurally different than a knife cut or had one or more tooth compression marks near one end, as well as associated pitting.
Carnivore chewing is evident on some foot bones which are normally difficult to break and would unlikely be broken by humans in the butchering process. The navicular bones MA 1715 shows edge crushing and MA 1668 has a section broken off. The calcaneus MA1709, MA1524, MA1826 and 1933 are crushed in half or have their end broken off. Most deer phalanges are smashed in half and two have channeling typical of carnivore chewed bones. MA1663 has distinct carnassial tooth marks inside the broken end.
Long bones that show carnivore gnawing characteristics such as chipping back of the edges and deep channeling at the centre of the shaft were not observed. Leg bone ends were proportionally more prevalent than bone splinters. This would suggest human rather than carnivore activity. Carnivores usually chew the soft ends and leave more of the central shaft portions.
The deer ribs are missing both ends suggesting they were chewed off rather than broken off. MA1471 has distinct tooth puncture marks on alternate sides of the rib toward the distal end.
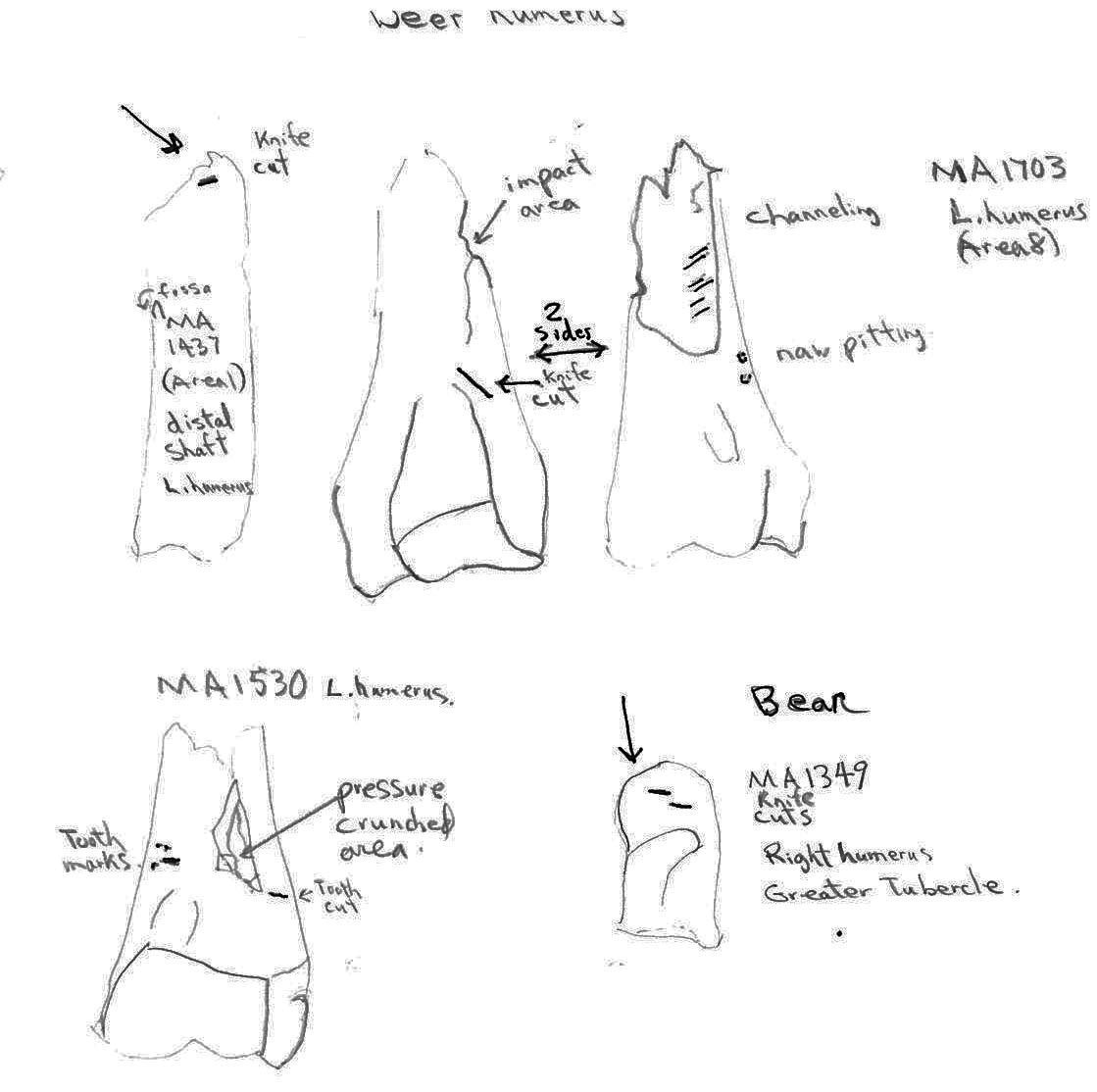
Two distal deer humerus show evidence of carnivore chewing. The distal end of humerus MA1530 has teeth cuts in line with a crushed area and humerus MA1703 has tooth pitting next to a channeled shaft. MA1703 also has a knife cut mark below the epiphysis. A deer radius MA 1400 has a tooth pressure impact on the distal end with knife cut marks under the epiphysis.
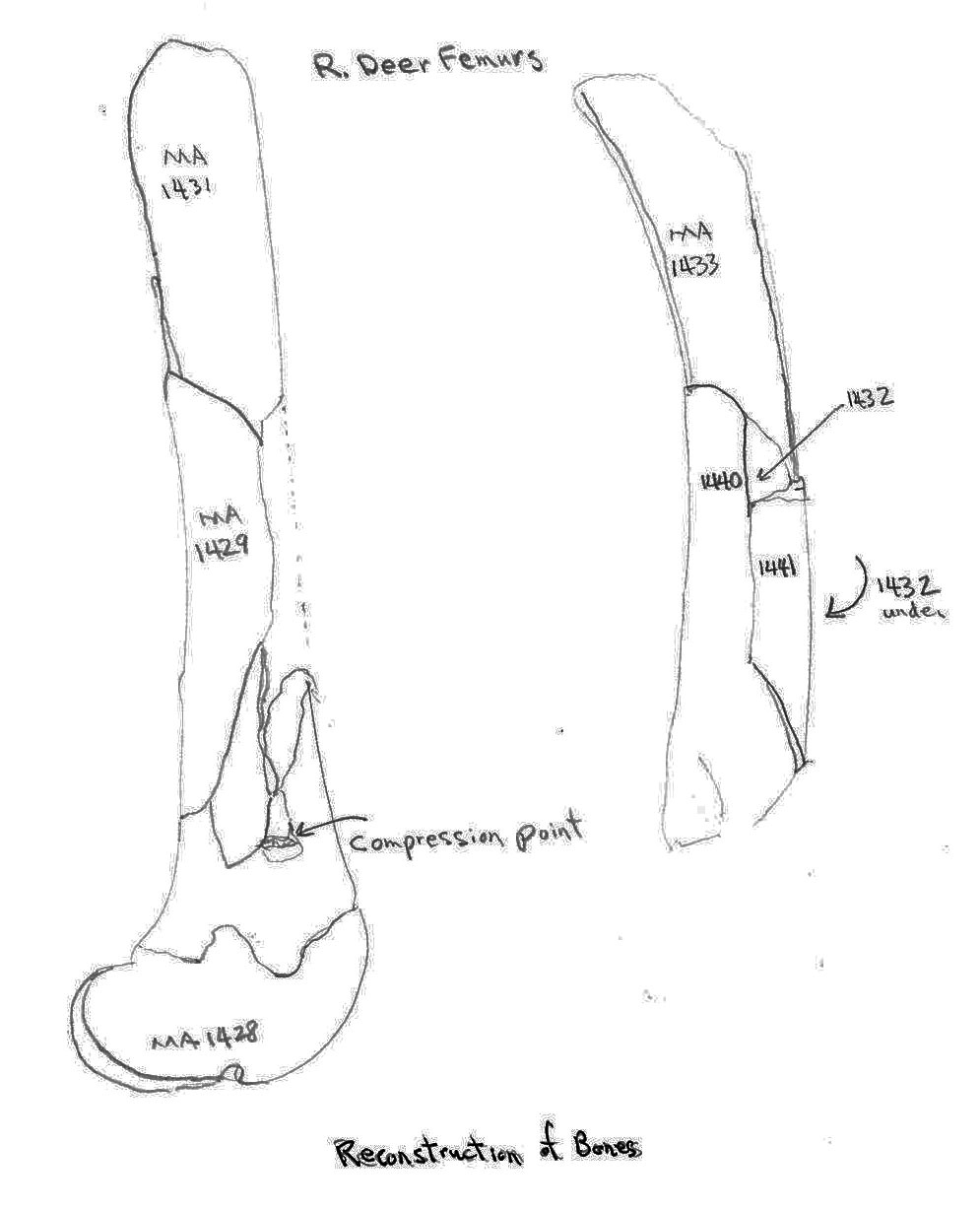
Two deer femurs were partially reconstructed from bone fragments. These showed no hammer-like impact fractures or knife cuts. One composed of bones MA1428, 1429 and 1431 shows a distinct tooth compression mark at the junction of the fractures.
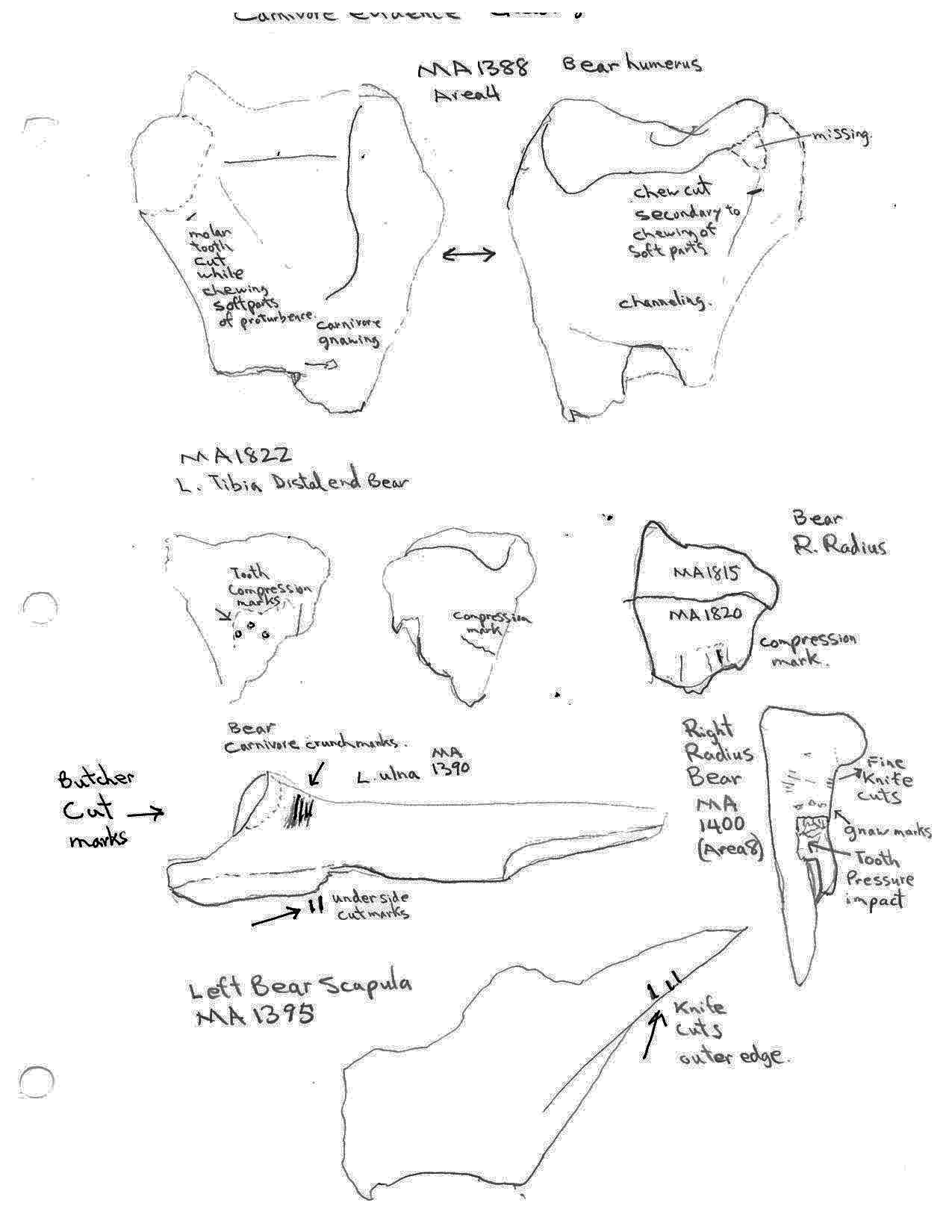
Carnivore chew marks are present on several bear bones. The distal humerus MA1388 has tooth cut marks on both sides of the bone adjacent to missing soft bone areas as well as gnawing and channeling below on the broken end of the shaft. A distal end of a tibia MA1822 and a distal end of a radius MA1815 have tooth compression marks. The proximal end fragment of a bear ulna has tooth cut marks on two opposing sides.
There is a high incidence of rodent chewing on all bones and a low incidence of carnivore chewing on the deer and bear remains. The marmot remains show no evidence of being chewed by carnivores. It cannot be determined if the carnivore marks were created by wild animals or by aboriginal dogs. Possibly dogs may have been present when the bear and deer were being butchered but not when the marmots were butchered. It would seem likely that marmot parts given to, or scavenged, by dogs would be consumed whole but this is not the case with some recent carnivore eaten marmot remains that clearly show only partial consumption of the cranium and mandibles.
Introduction To The Study Of Human Bone Modification
As a general rule knife cut marks derive from different stages of processing an animal. Some cuts are indicative of skinning, some of primary butchering, and some of secondary butchering.
Human induced butchering and skinning marks should be expected to show two main criteria. (1) Repetition of cut marks on many examples of the same bone element. (2) Some anatomically dictated reason why a particular cut mark should occur at a given location on the bone.
In skinning there are few places on the anatomy where the removal of the skin brings the butcher’s knife in direct contact with bone. The two places where this is most likely are the lower legs and the head.
Skinning often results in cuts across the phalanges, the lower tibia or the shaft of the metatarsal, around the base of antlers, ears, the sides of the nose, and around the chin area of the mandible and maxillae.
In butchering, or the disarticulation of the skeleton, cuts generally occur around the base of the skull or on the first two cervical vertebrae. Cuts occur on or near the articular surfaces of the long bones where ligaments and muscles need to be cut. Filleting of the tenderloin and the removal of rib slabs may result in cut marks on the body or spinal processes of the vertebrae, cuts on the surface of pelvic parts or on the middle or end of ribs.
Characteristics Of Cut Marks
Cuts from tools are deeper on the parts of the short curvature and shallow on the flatter or long curvature surfaces. This is different from scoring by carnivores which tend to follow the contour of the bone. Carnivore scoring is often transverse to the long axis of bones since it results from turning the bones against the teeth. There may be tooth scores parallel and close together which superficially resemble a series of knife cut marks. Carnivore tooth scoring also occurs on flat surfaces next to a chewed edge.
Hammering or smashing of bone can be expected in the case of larger animal bones for the extraction of marrow; or for acquiring bone sections or splinters for raw material in tool manufacture. Canines may be extracted for use as body adornment.
One objective of this study will be to try and reconstruct the skinning pattern and sequence of animal carcass disarticulation after skinning. For example, butchering often takes place by part removal in the following sequence: 1. Cranium 2. Front legs, clavicle, scapula. 3. Rear legs, pelvis. 4. Basic axial skeleton-vertebral column, ribs.
The Human Alteration Of Marmot Bones From Clayquot Cave

Modification Of The Cranium
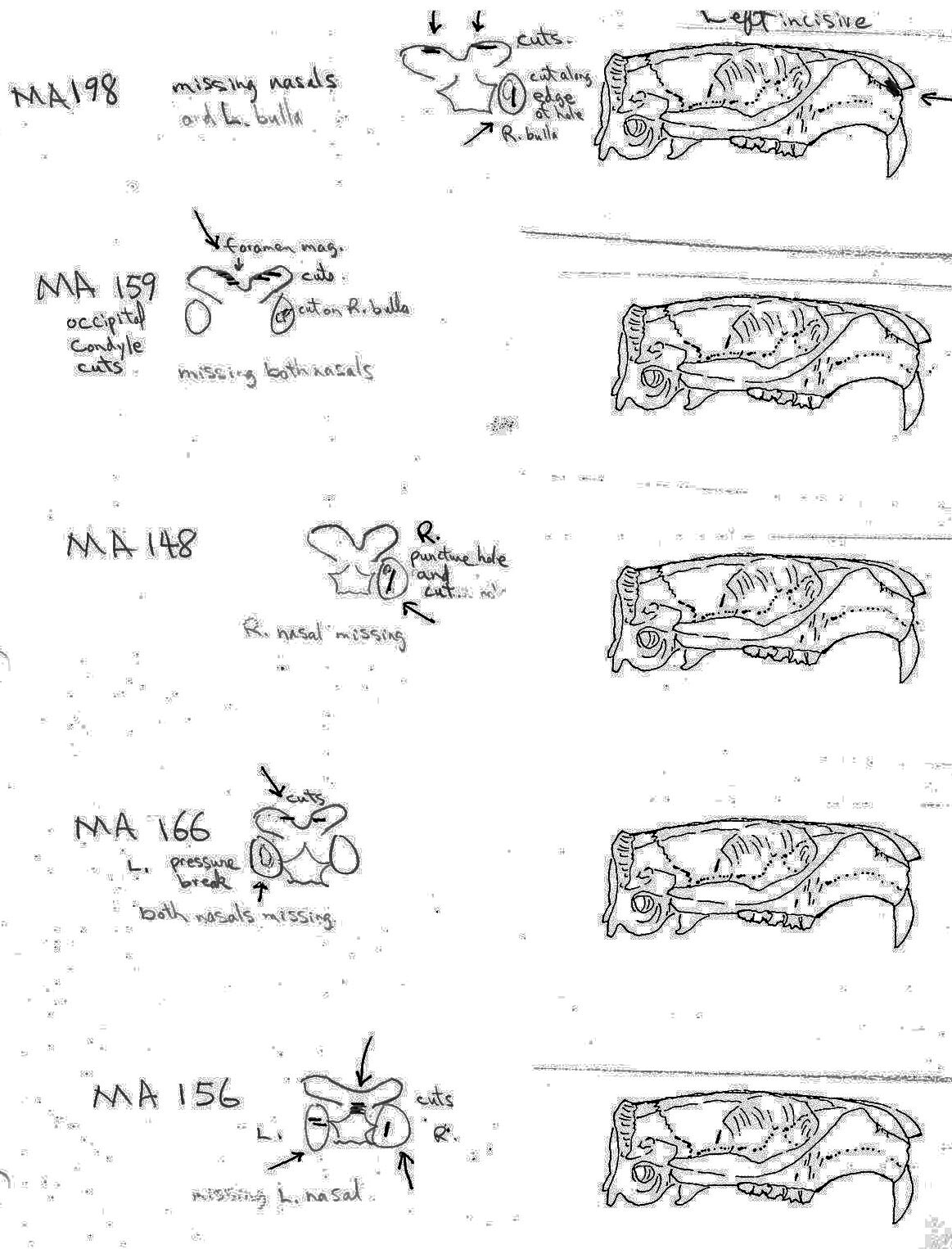
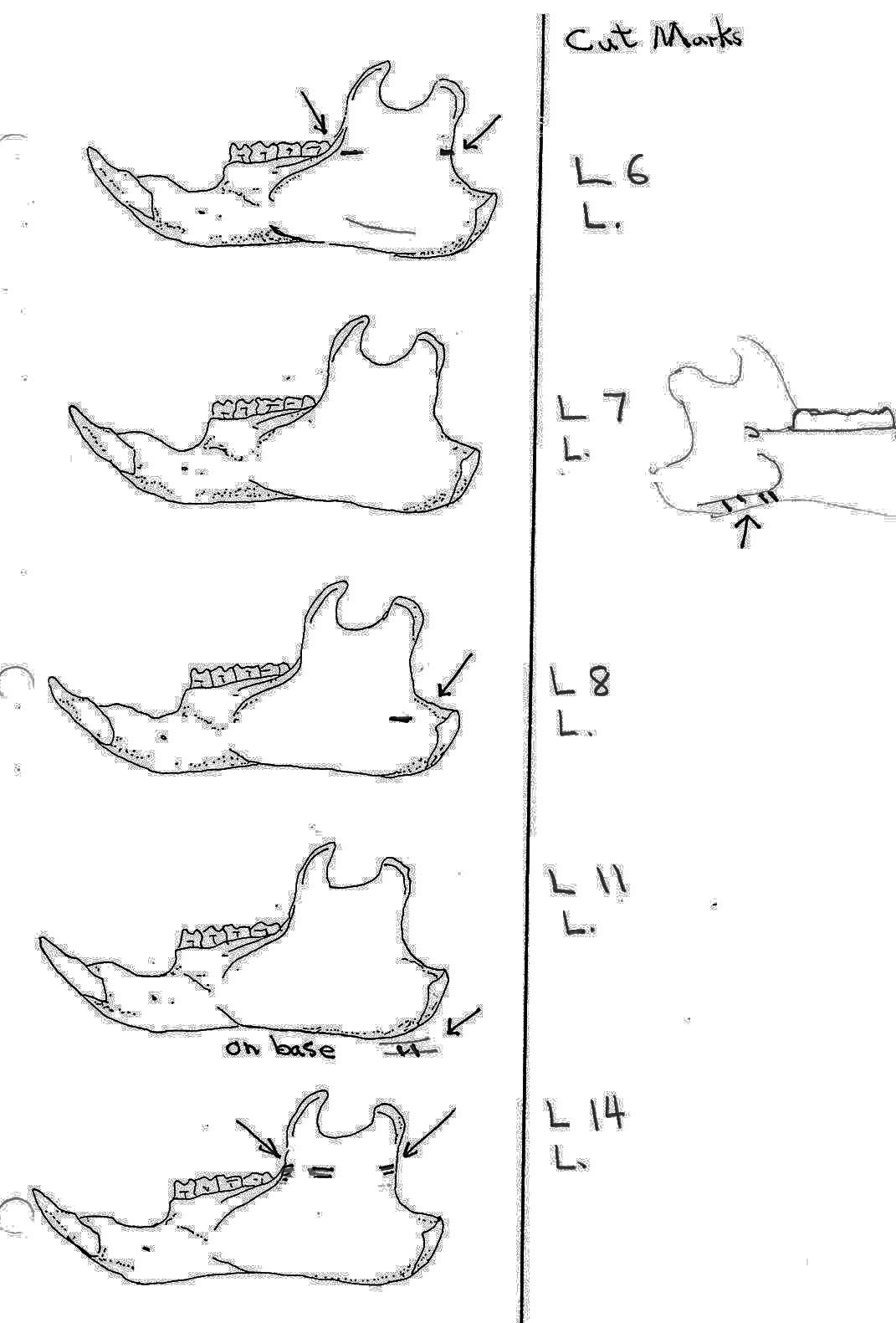
The MNI from this site is 14. Eleven of these are represented by the cranium, of which 9 show evidence of work marks on the distal anterior only. One specimen that is unworked is missing its anterior and posterior end. Most cuts are on the incisive bones. Four have right incisive cuts only, 2 have left incisive cuts and one specimen has cuts on both incisives. A premaxillae fragment also has a left incisive cut. One right and one left nasal bone are cut and one specimen has a long cut on the top of the left frontal in addition to right incisive cuts.
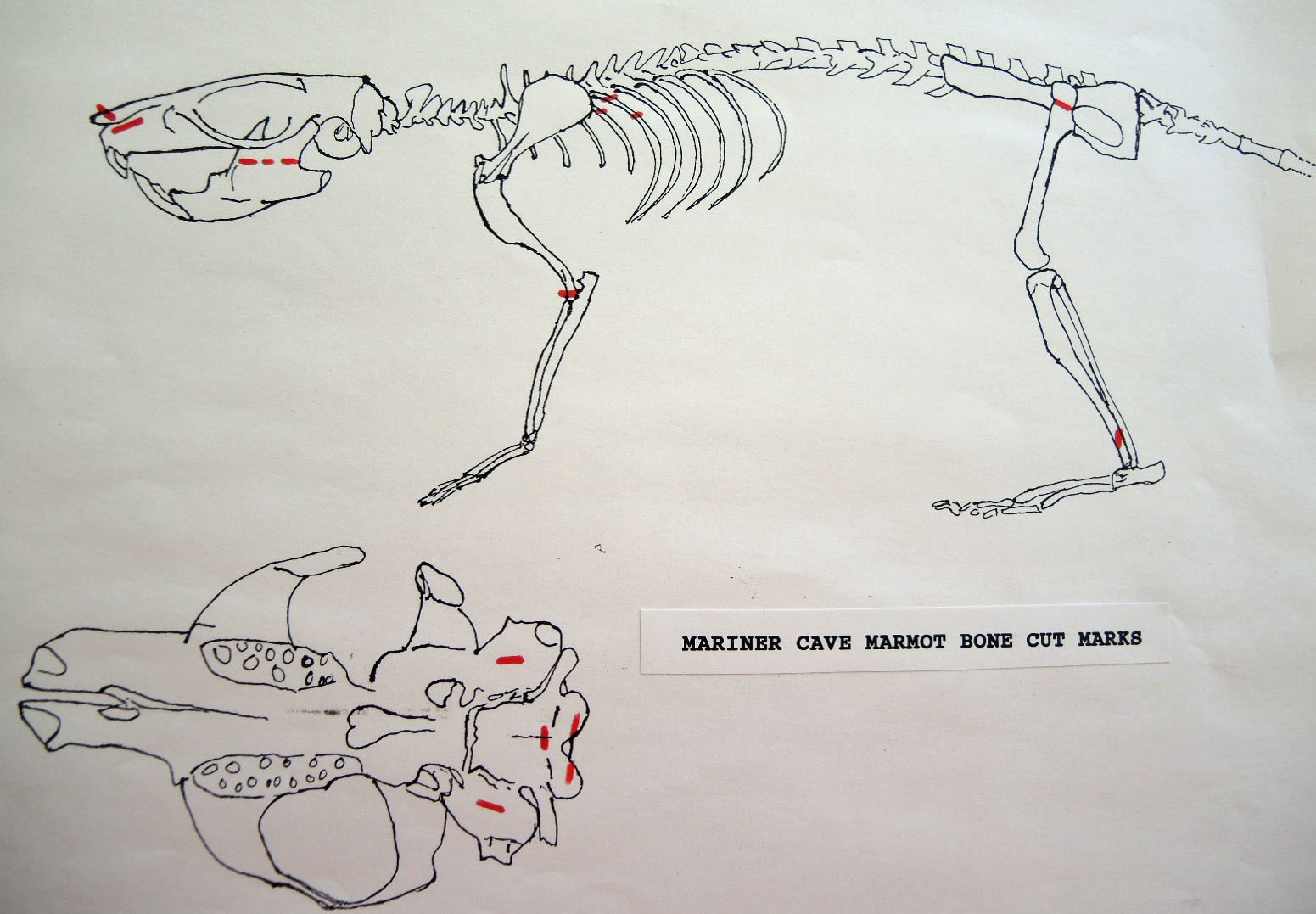
Evidence shows that the head of the animal was snapped forward to remove it rather than being severed near the base. All of the cranium have been broken close to the occipital suture line. The occipital are only present for 6 individuals, the others are missing. Seven auditory bullae have puncture marks that are consistent with being punctured by the pterygoid at the base of the skull when the cranium was snapped forward.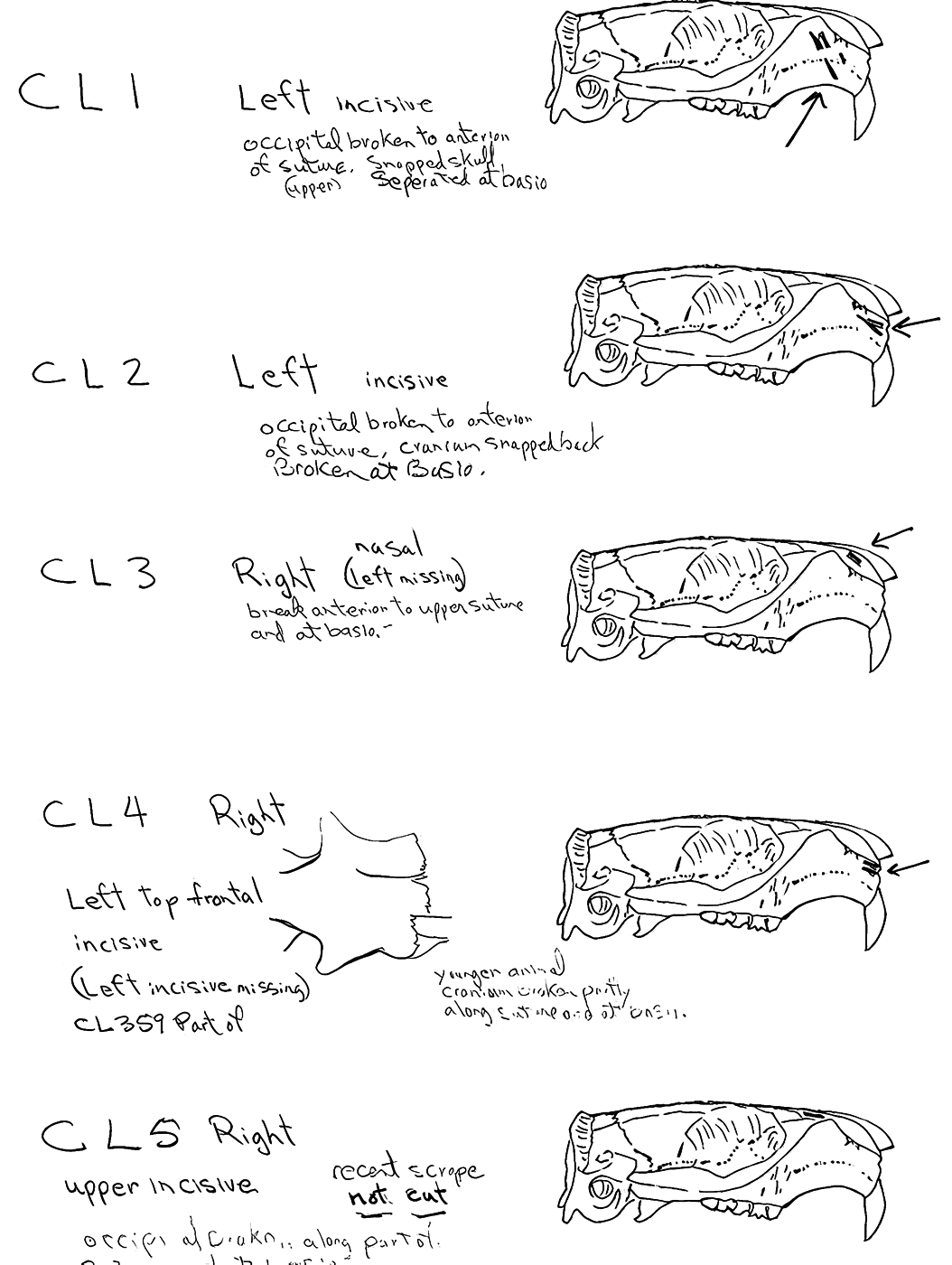
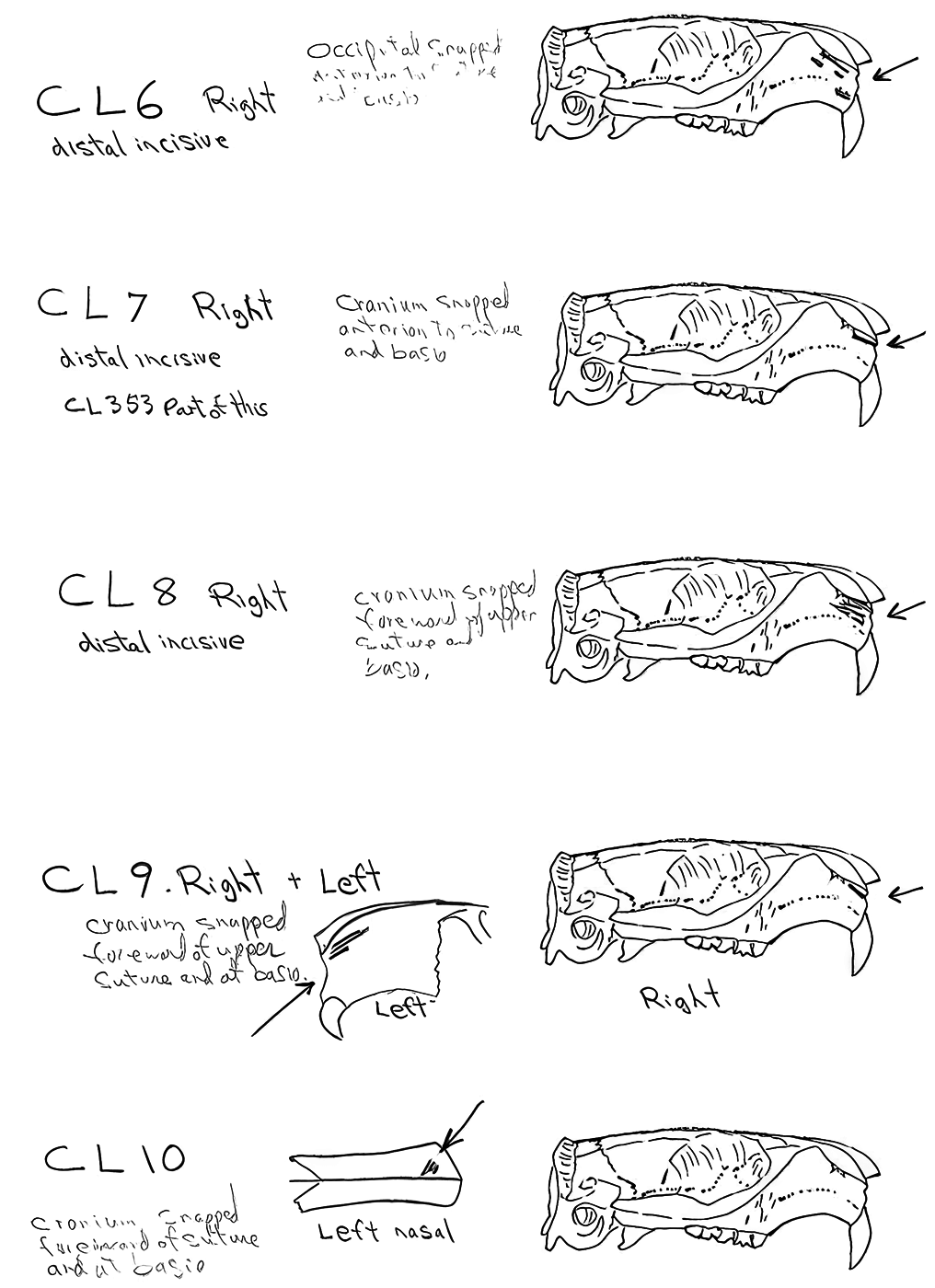
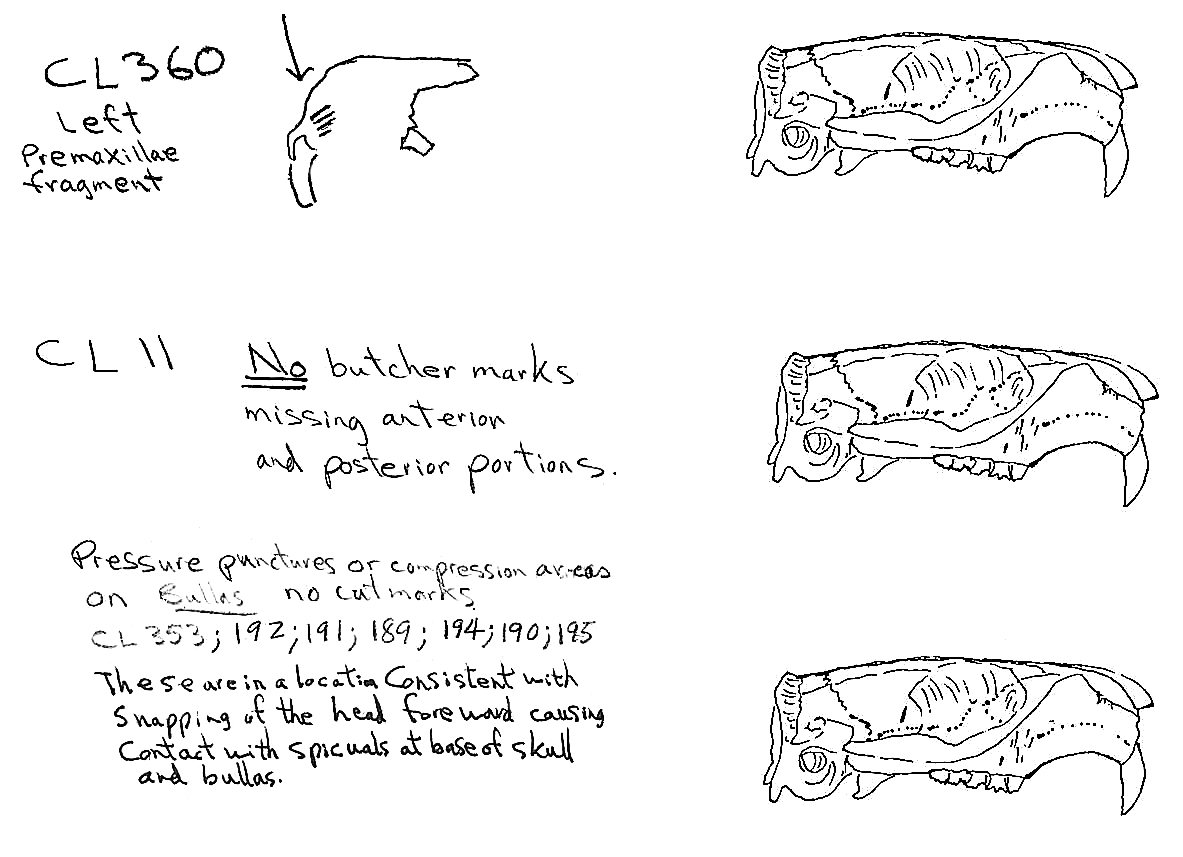
Modification Of The Mandibles
A minimum of 14 individuals are represented by mandibles. Six of these are left mandibles and have skinning cut marks on the anterior portion of the ramus. One right specimen has three cuts under the base of the posterior ramus which likely results from skinning or cutting of the throat area before the head was snapped back.
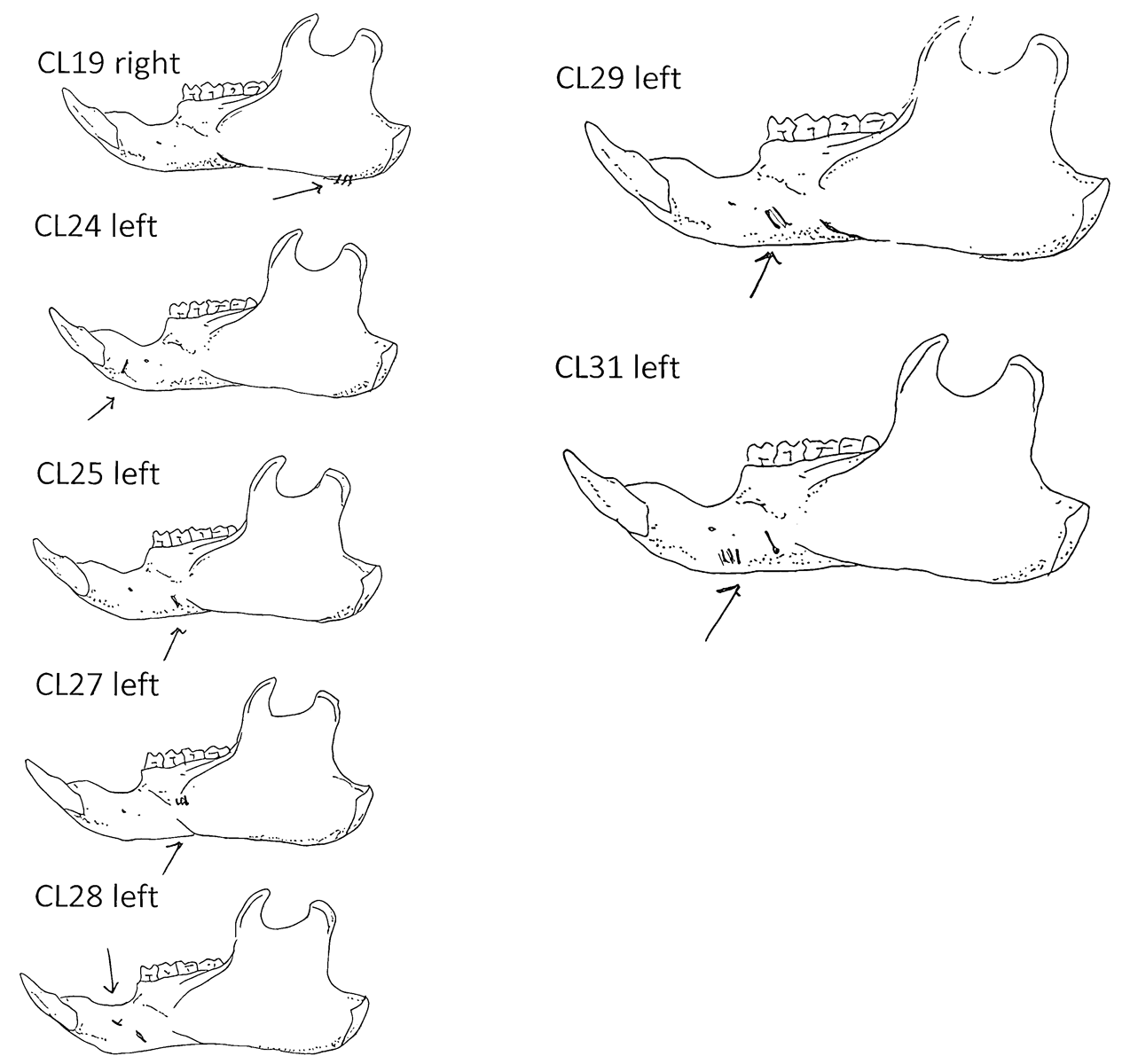
Modification Of The Femur
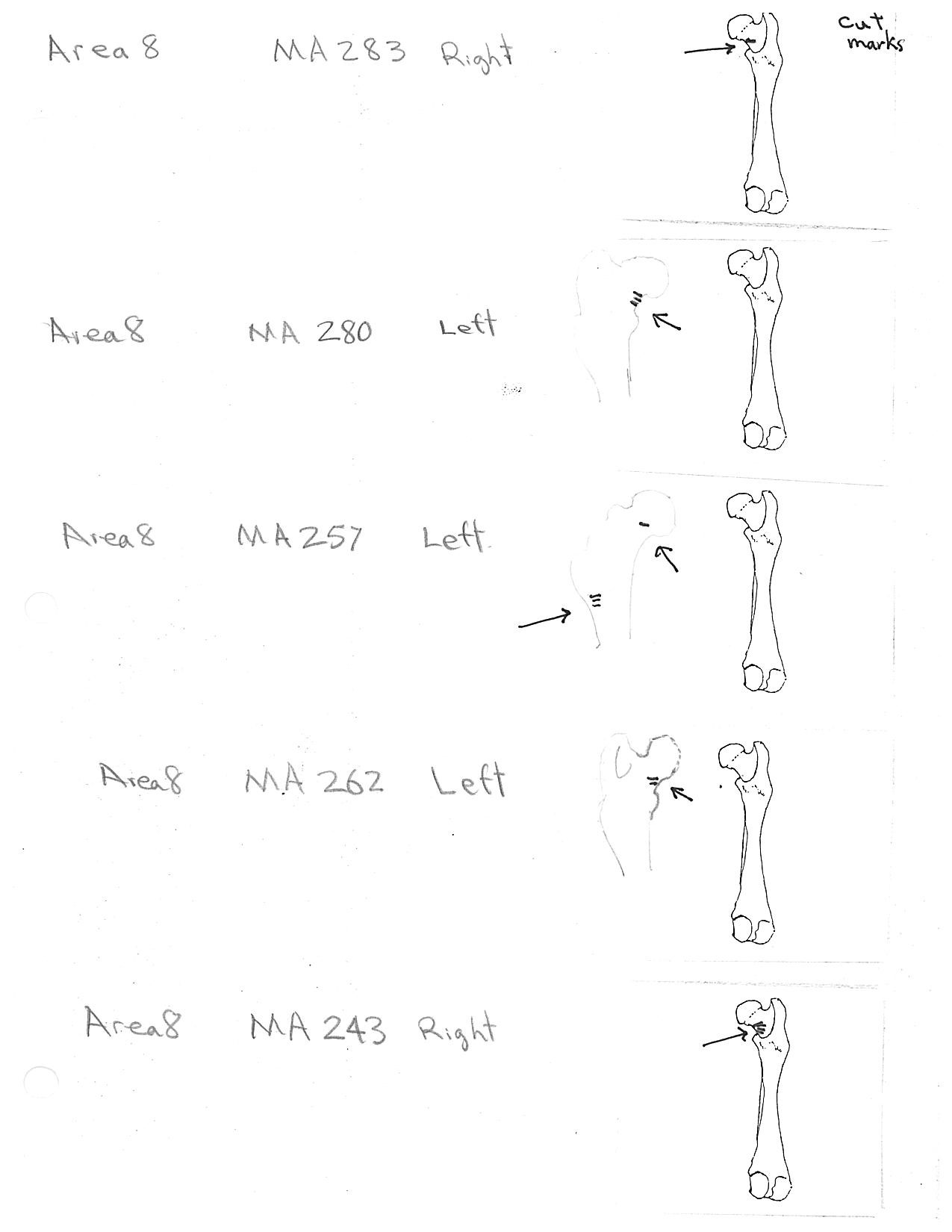
Ten femurs have been cut. All ten have skinning cut marks on or just above the distal end on the posterior side. Two of these also have butcher marks on the femur neck.
Modification Of The Humerus
Five right humeri have cuts near their distal ends. Two are cut across the bone bridge above the lateral epicondyle and three have cuts at an angle to the long axis of the bone near the distal end of the shaft.
Modification Of The Clavicle
Eight of the ten clavicles from the site are cut. Four right and four left clavicles show cuts mostly in mid shaft or toward the distal end. Only one is cut near the proximal end. These are likely butcher marks intended to cut the shoulder muscles.
Modification Of The Metatarsals And Ribs
Two right metatarsal have two cuts each across the surface of the bone. There are only three ribs from the site. All have broken distal ends and only one has two sharp cuts across its midsection.
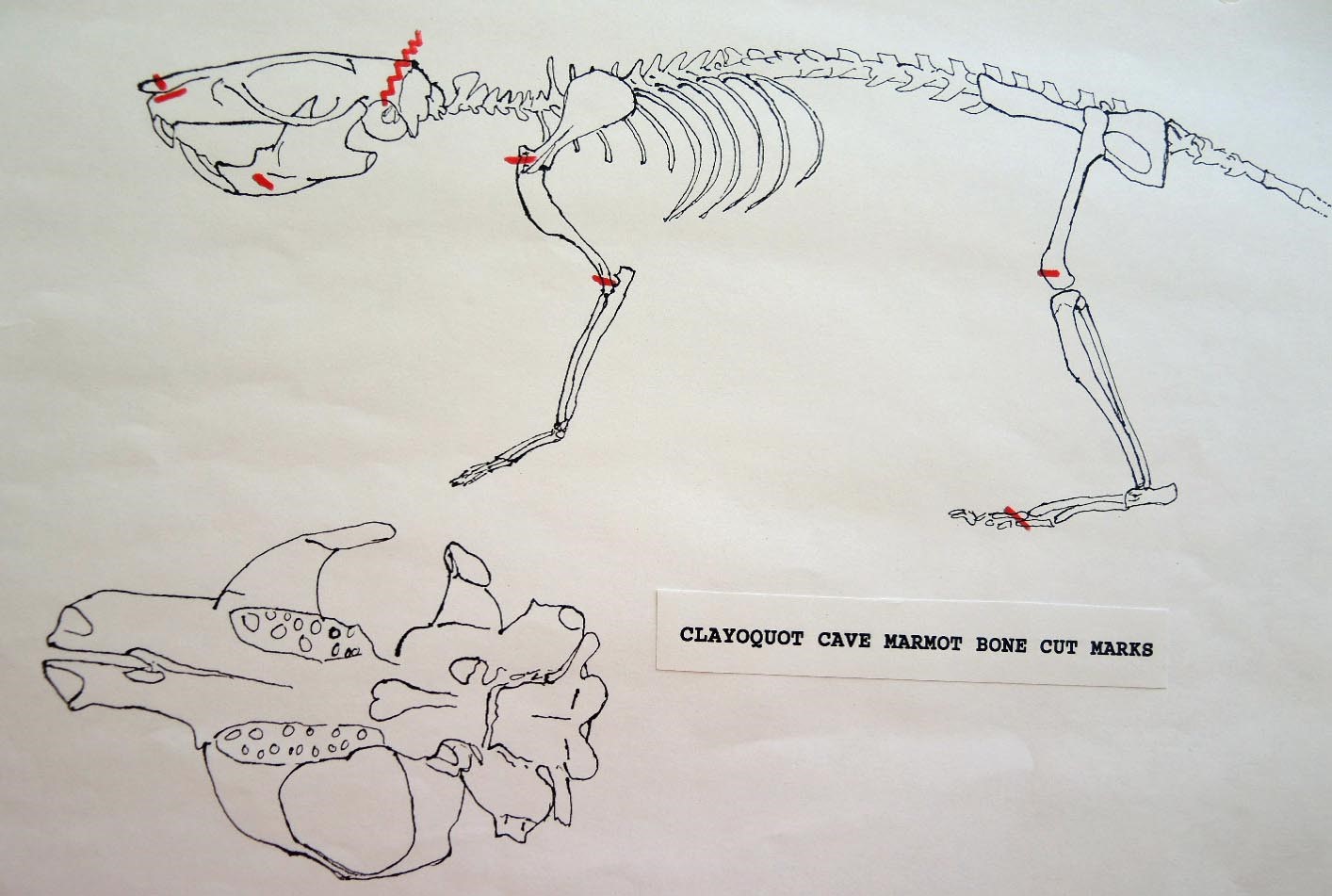
Marmot Bones From Clayquot Cave
SUMMARY LIST OF PRESENTS AND ABSENCE OF CUT MARKS
(MNI = Minimum number of individuals)
(Showing MNI represented by each cut element class and the total MNI in the site represented by that element)
| BONE ELEMENT: | NUMBER WITH CUT MARK(S) |
|---|---|
| Cranium | 9 (MNI 9) (site 11) |
| Mandible | 7 (MNI 6) (site 14) |
| Femur | 10 (MNI 5) (site 11) |
| Humerus | 5 (MNI 5) (site 13) |
| Clavicle | 8 (MNI 4) (site 6) |
| Ribs | 1 (MNI 1) (site 1) |
| Metatarsal | 2 (MNI 1) (site 3) |
| Tibia | 1 (MNI 1) (site 13) |
Bone elements with no cut marks: Fibula, scapula, ulna, radius, innominate.
Bone Modifications From Clayquot Cave – Conclusions
The marmots were skinned across the upper maxillae where cut marks are evident and possibly across the face to the top of the neck based on cuts on the posterior underside of some jaws. The head was then snapped forward often breaking at or near the occipital suture line and causing the pterygoids to puncture the auditory bullae. There is no evidence of the cutting off of the tongue, the cutting of the cranium at the base or the cutting of the masseter muscle attached to the side of the skull.
Skinning occurred by cutting into or around the distal end of the femur and the humerus. The general absence of foot bones and a few metatarsal cuts indicate that the feet were left on the pelt.
Only a few femurs were cut off at the pelvis, the others may have been separated with the abdominal area when it was snapped and pulled off. Cuts on the clavicle suggest butchering of the forelimb section by cutting the shoulder muscles to the clavicle. The uncut humerus and scapula were part of the pulled off forelimb section.
At least part of the rib cage of one animal shows butchering marks. This is likely an unintentional secondary effect of cutting into the rib cage when removing the upper forearm section.
The Human Alteration Of Marmot Bones From Mariner Cave

Modification Of The Cranium
The minimum number of individuals based on skulls is 62. This includes fragmentary remains that can not be accounted for by missing parts of the more complete specimens. That is, they overlapping with existing large parts of the cranium. In the case of maxillae fragments, which represent 9 different individuals, only two could possibly match missing sections of skulls and therefore represent 7 individuals.
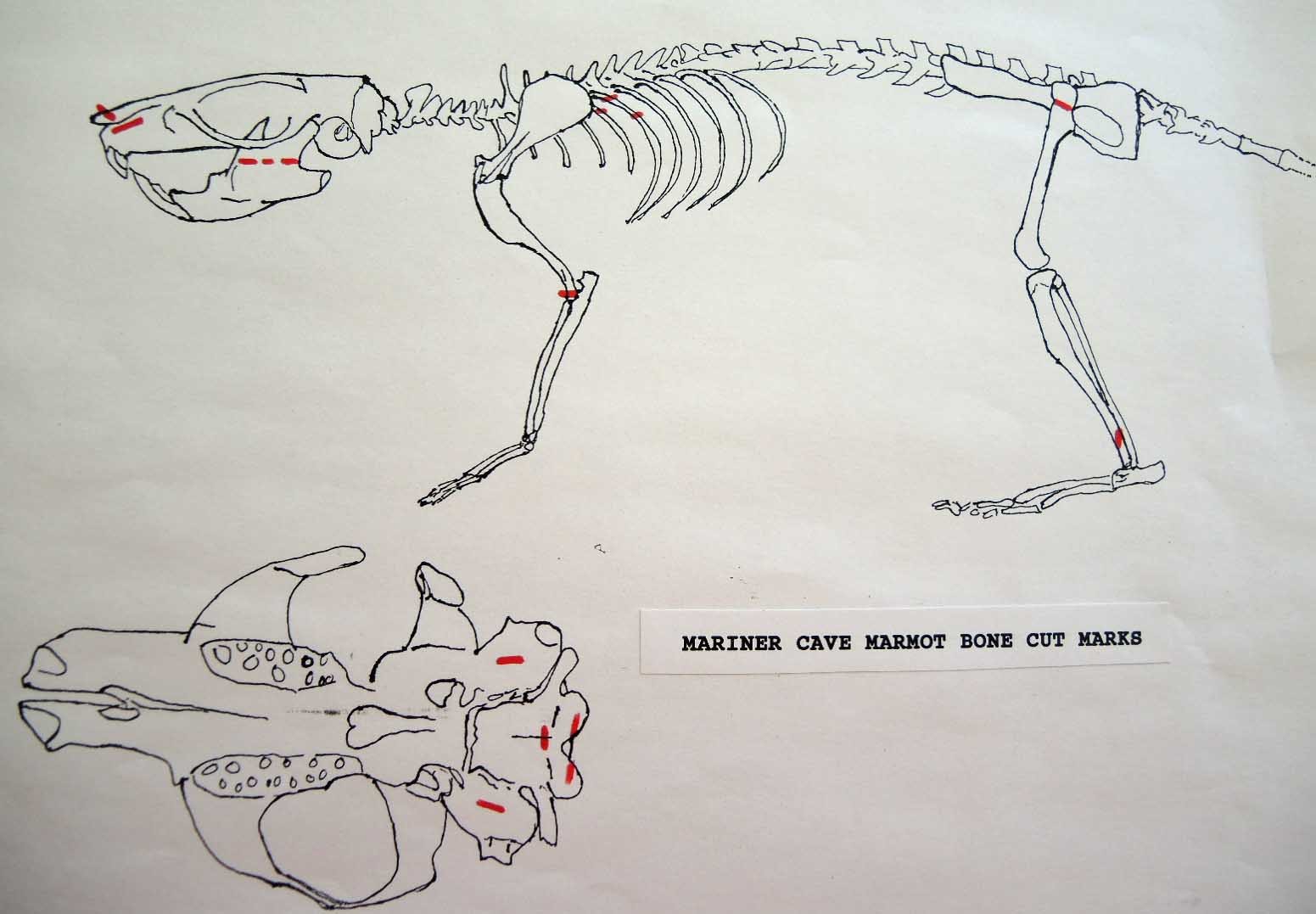
Cut marks are found on skulls representing 23 individual marmots as well as being found on an additional 28 separate skull parts which cannot be proven to be exclusive of the 23. These 28 separate parts in themselves represent a minimum of 8 individuals. Of the skulls with no cut marks (MNI 39), only 7 have all the parts that normally have cut marks. Of those skulls representing individuals with cut marks, all but one have parts missing which may exhibit further cut marks.
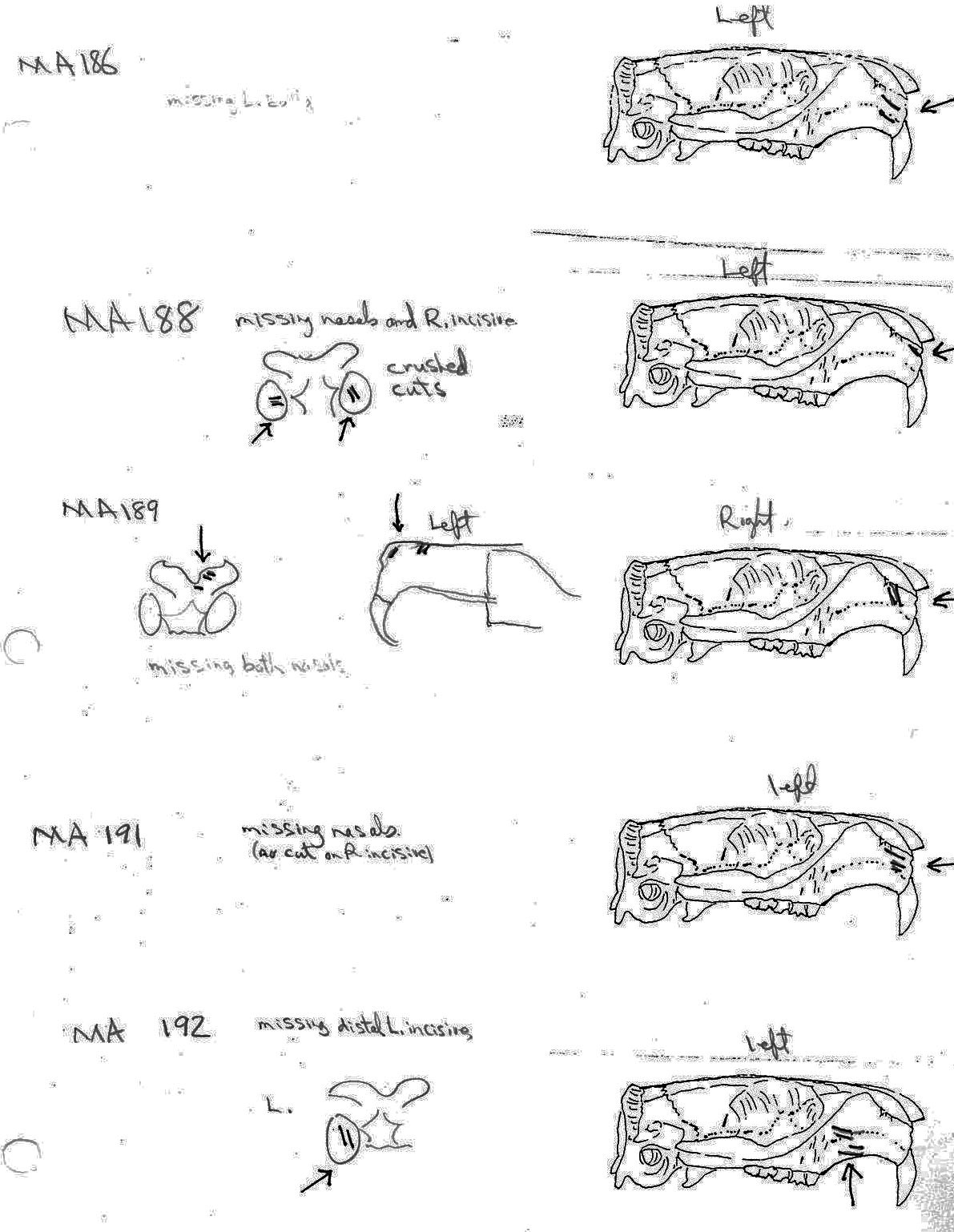
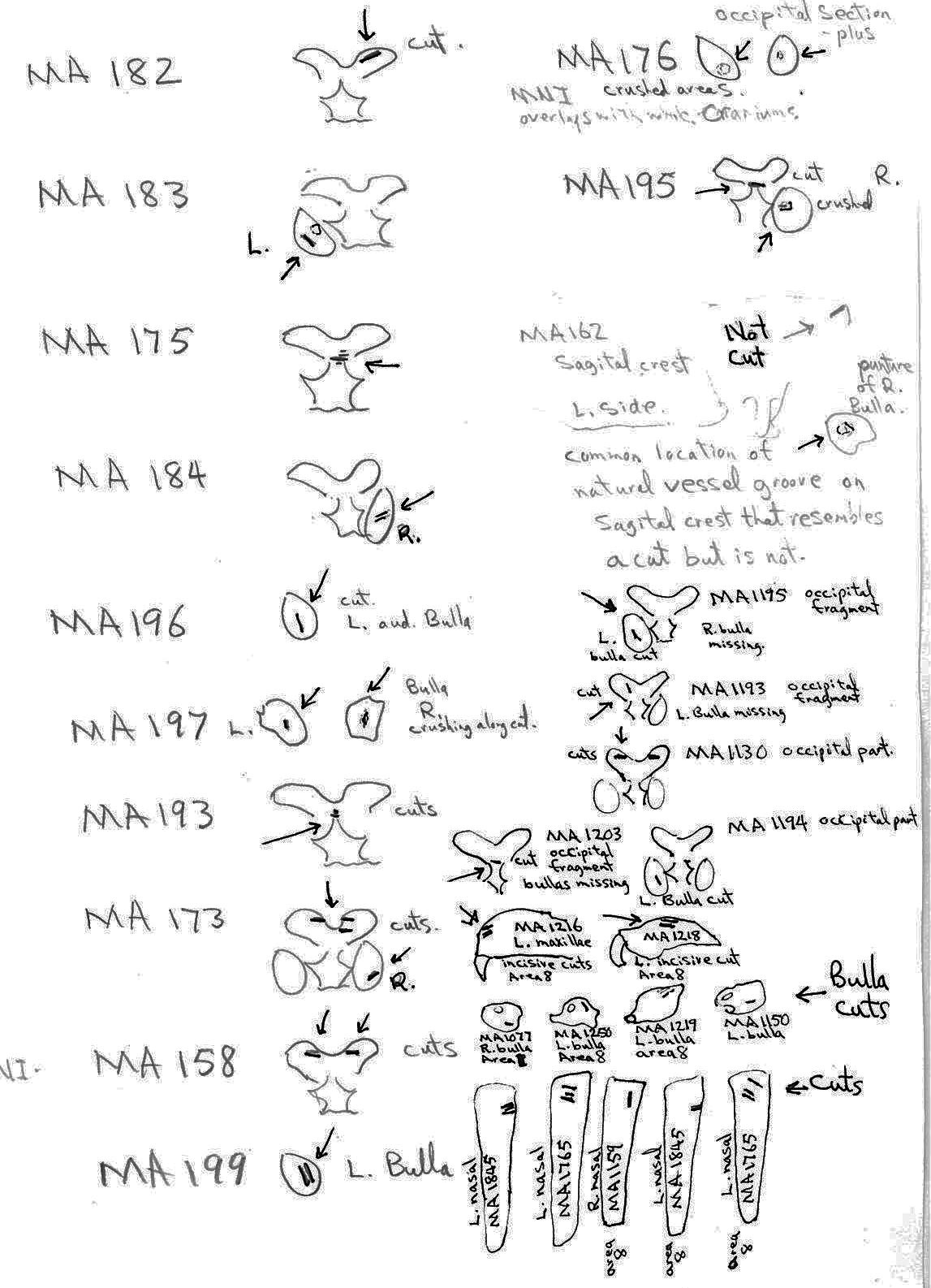
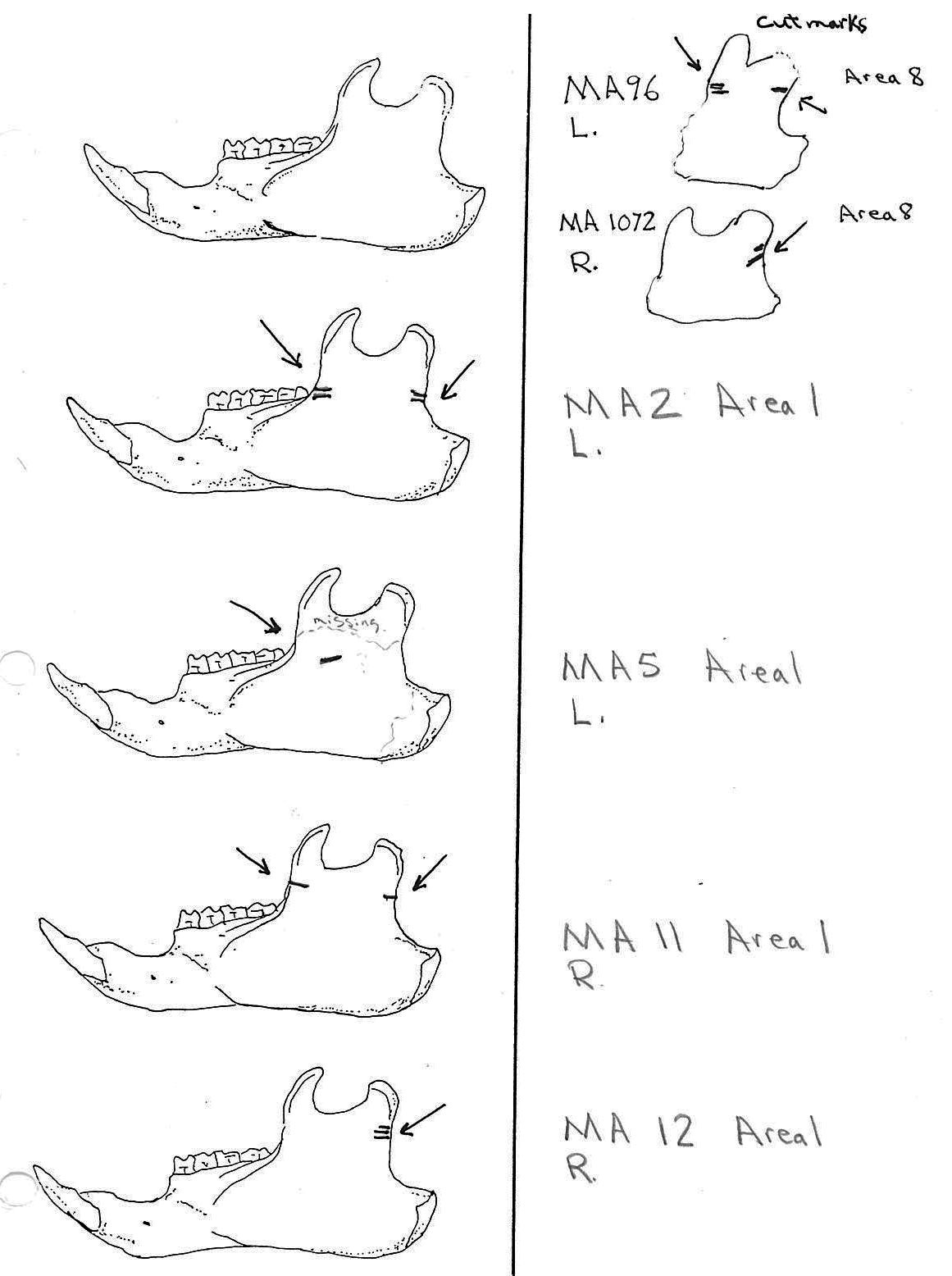
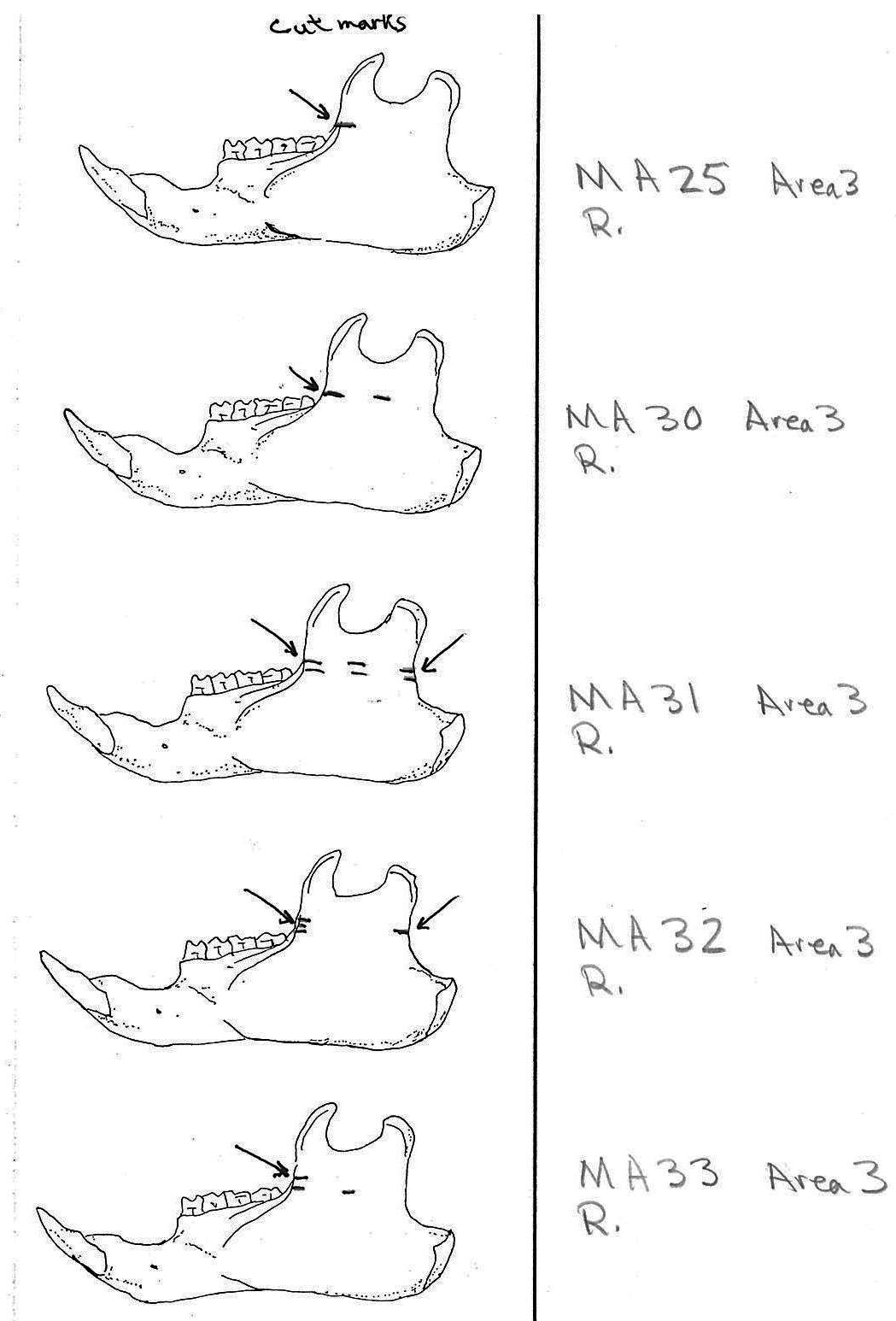
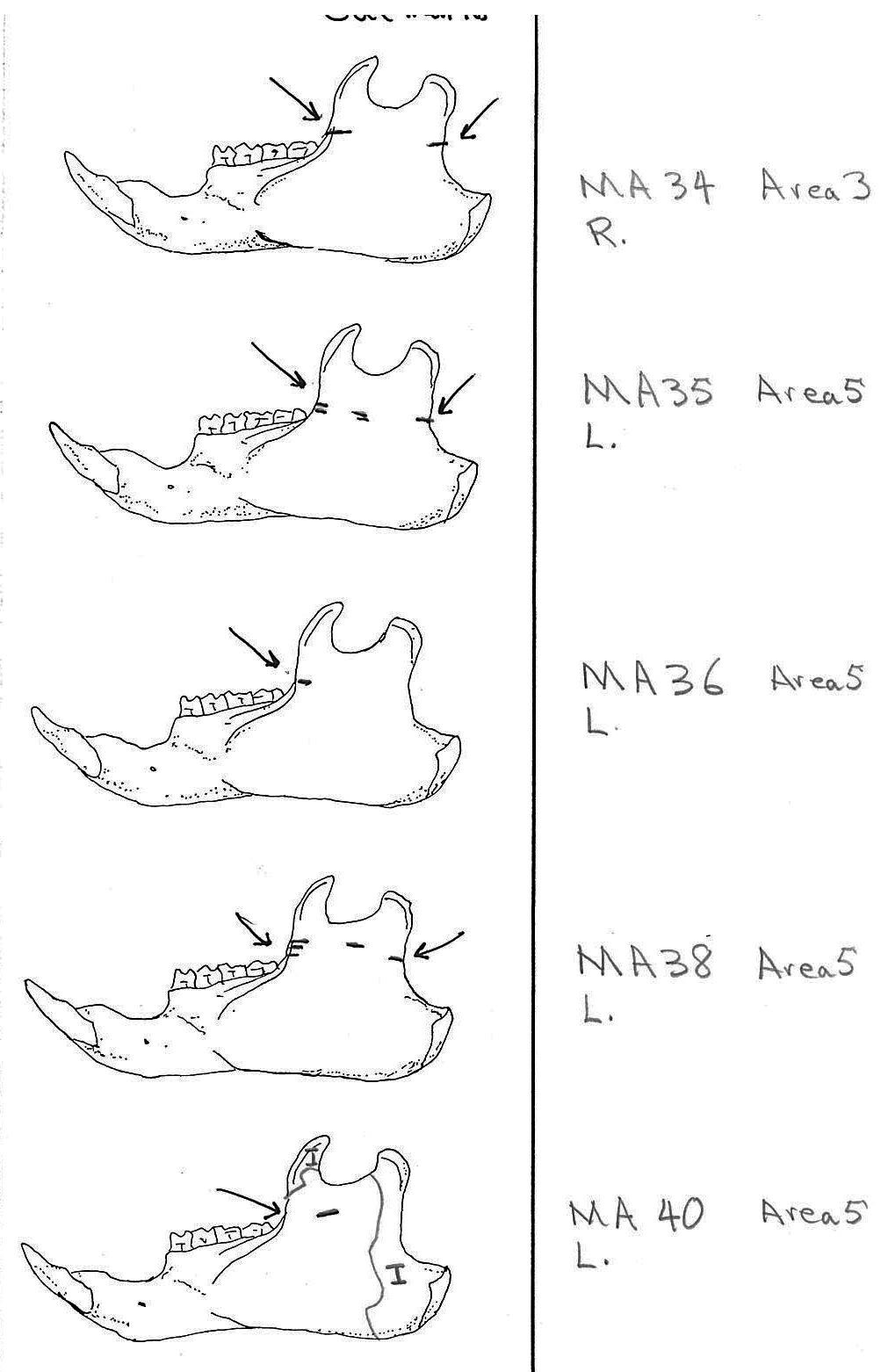
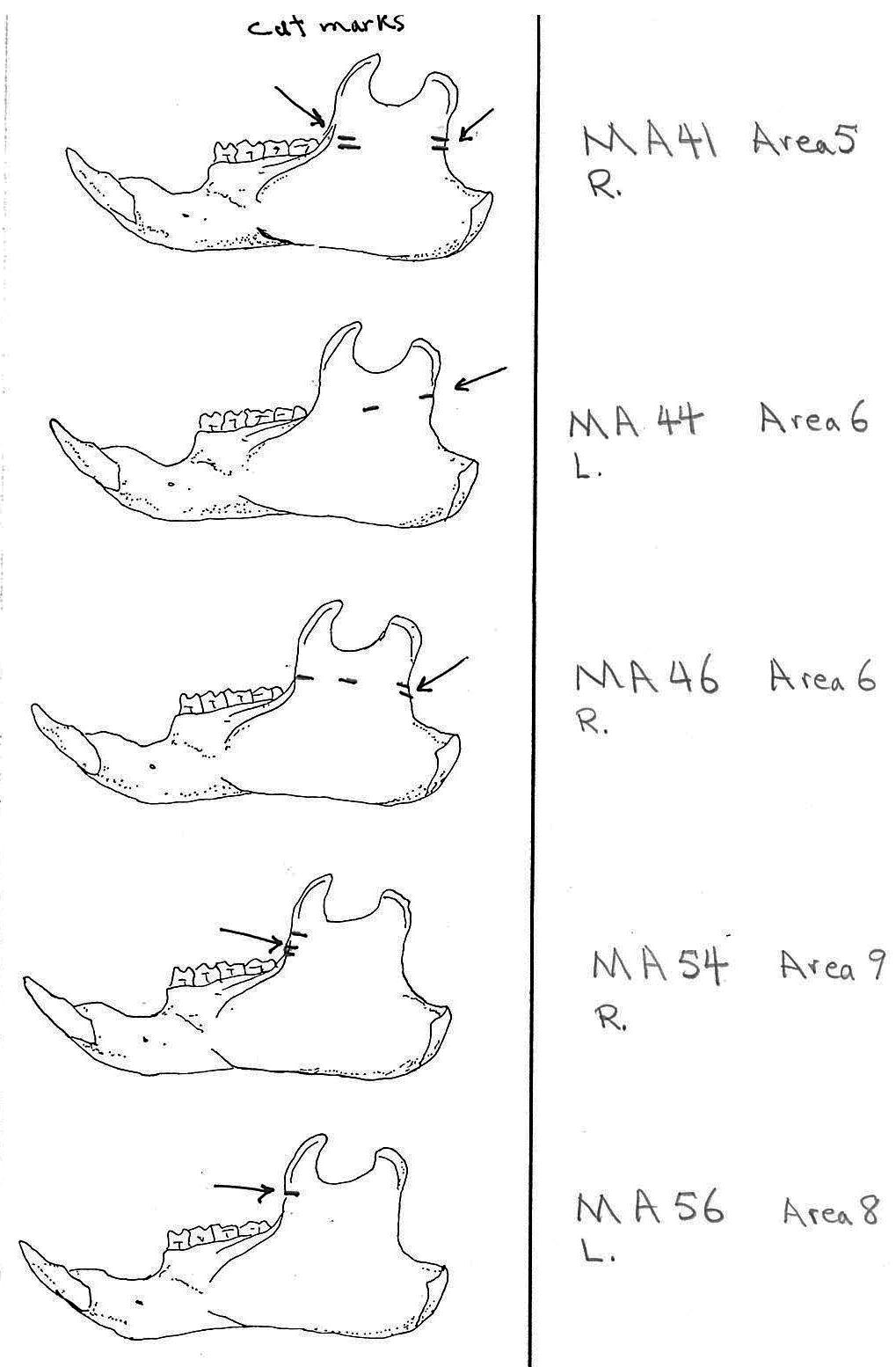
All cuts on the skull are located on the upper anterior of the maxilla and on the ventral occipital in various combinations. Cut locations include 16 left and 5 right incisive bones. The cuts are usually along the upper anterior edge of the incisive which would correspond with the area just below the sides of the nose. Cuts also occur on the anterior portions of 4 left and 1 right nasal bone. This corresponds to the area next to the top of the nose. All of these maxillae cuts are indicative of skinning.
Cuts occur across the articular surfaces of 7 occipital condyles and in 5 cases across the small ridge extending from the anterior of the condyle to the basio-occipital. No combinations of the latter two cut locations were observed. The cut angles on the condyles show that the cutting was directly from the side of the skull posterior to the bullae. These cuts are typical of those induced when the head is pulled back and severed from the neck.
Cuts, small crushed points, and combinations of these occur on the surfaces of the bullae. These include cuts on 17 left and 16 right bullae. Seven of the latter are matched pairs. The positioning and angle of these cuts and punctures suggest that a knife blade point was jabbed into this area to sever the muscles which attach the back of the tongue to the skull.
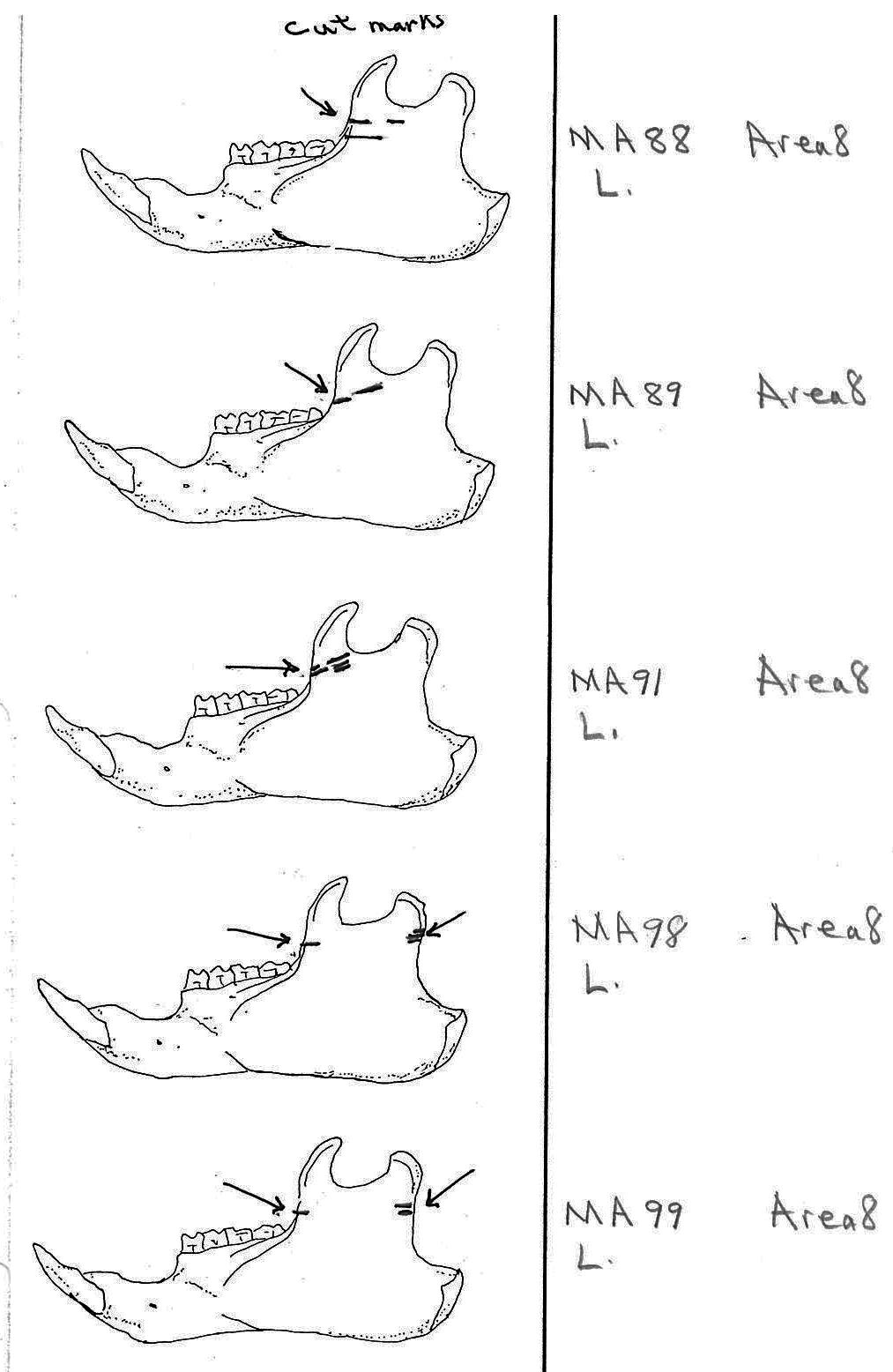
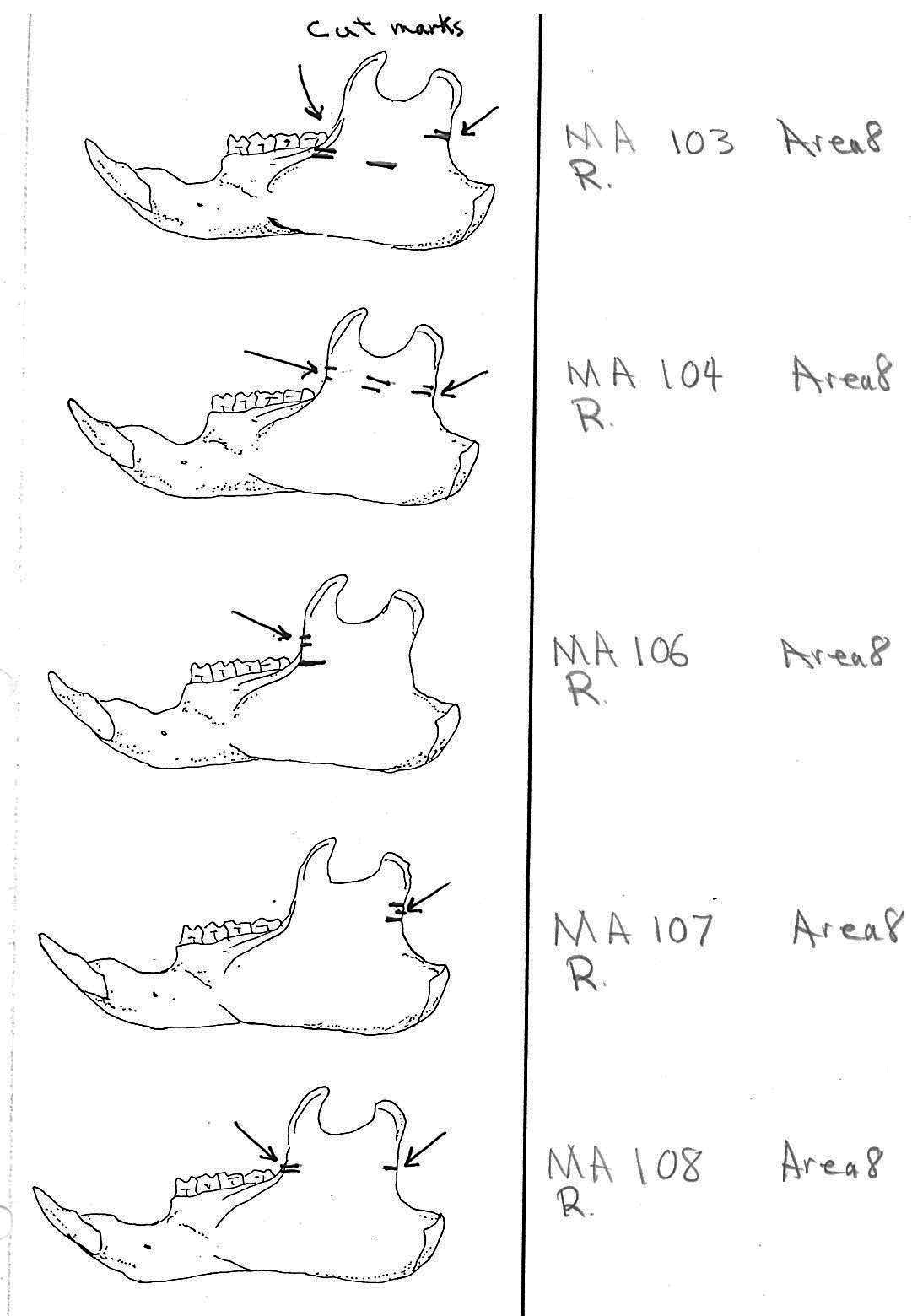
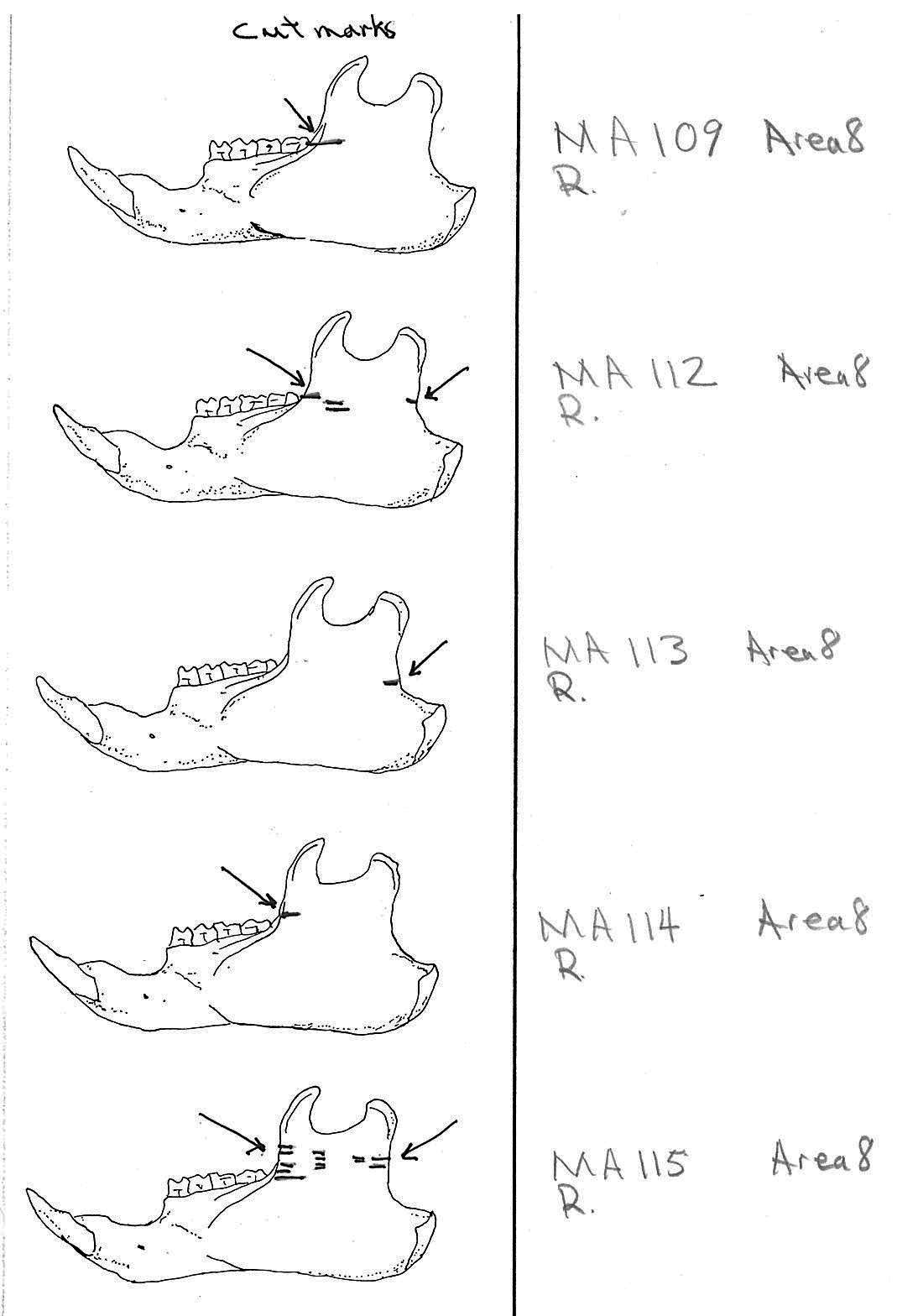
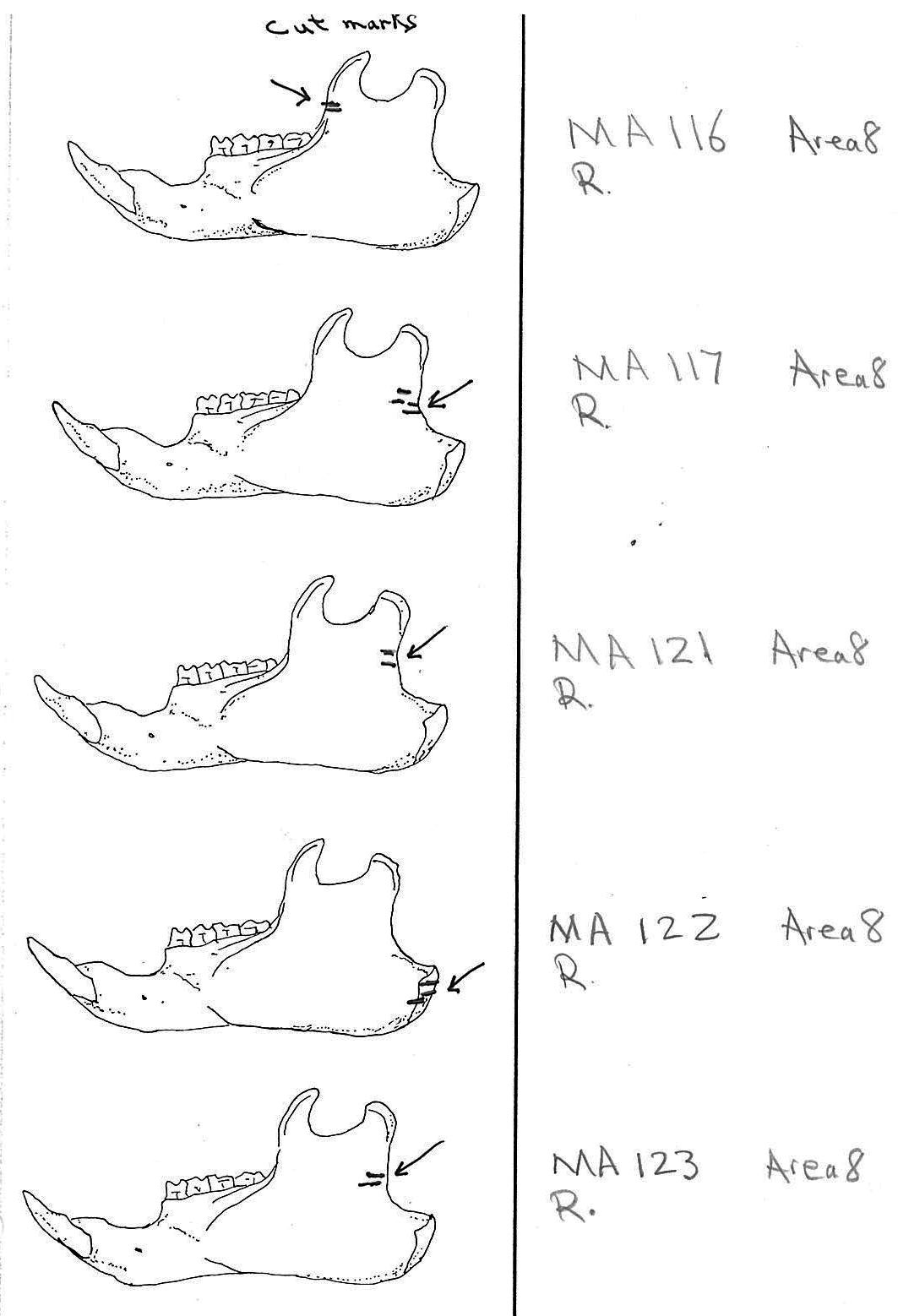
Modification Of The Mandibles
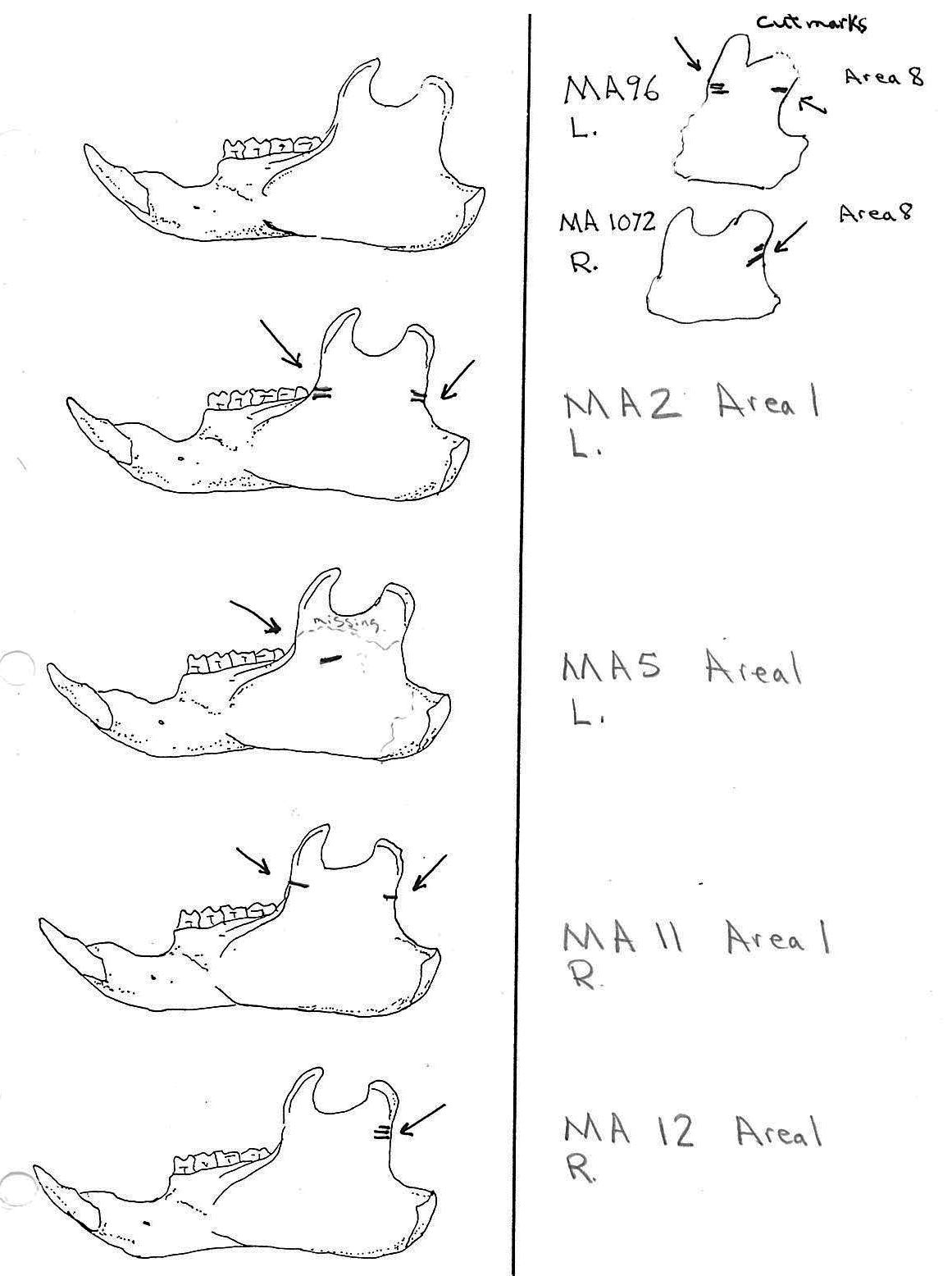
There are a minimum of 74 marmots represented by left mandibles. Right mandibles number 73. Cuts occur on 41 right mandibles and 39 left mandibles. Incomplete or corroded parts where cut marks often occur are found on 36 specimens. Of the 31 complete specimens that have no cut marks, 9 are small mandibles of younger individuals. This suggests a slight bias toward fewer cut marks on younger individuals due to reduced effort in muscle cutting. However, distinct cut marks do occur on some of the smallest specimens.
Of the 80 mandibles with cut marks, all but 3 have the cut locations across the ascending ramus. As the skin in the ramus area is loose and easy to cut it is not likely that these marks would be made during the skinning process. Only when cutting deep to remove the jaw by severing the masseter (pars caudalis profunda) muscle below the zygomatic arch would these kinds of marks be produced.
The cuts occur in some cases only near the edge at the base of the coronoid crest or front of the ramus (17 specimens); only near the edge on the posterior base of the condyloid process or back of the ascending ramus (11 specimens); across the centre of the ramus (4 specimens); or, more commonly, in combinations which include the first two cuts mentioned on both sides of the ascending ramus (22 specimens); and the latter combination plus other cuts near the centre of the ramus (15 specimens). Less common are combinations near the front and centre of the ramus (5 specimens) and the back and centre or the ramus (3 specimens).
In most cases the direction of the cutting is straight across the ascending ramus. In a few cases (e.g. MA89; MA91) the cuts angle up sharply from the base of the coronoid crest toward the centre of the notch between the coronoid and condyloid process or (MA57) from the centre of the ramus to the upper posterior of the condyloid process. The combination cuts across the ramus are often parallel, suggesting single knife movements across the ramus. Other cuts suggest separate cutting movements down near each edge of the ramus, but these are usually in the same horizontal plane. Cut angles across the ramus are substantially different in only one case (MA68), where the front cut is horizontal to the ramus body and the posterior cut is upward at a sharp angle across toward the back of the condyloid process.
Two mandibles have single cuts only on the base of the horizontal ramus curving under the masseteric line. These are likely the result of skinning the facial fur. One specimen has three cuts across the posterior edge of the angular process. This is likely to be an unintentional cut made when the knife came into contact with the back of the jaw during the severing of the cranium. Only one mandible, which has typical cutting marks on the ascending ramus, also has a single cut on the labial side of the mandible along the horizontal ramus. This is likely a random cut caused by the knife when cutting the muscles at the back of the tongue.
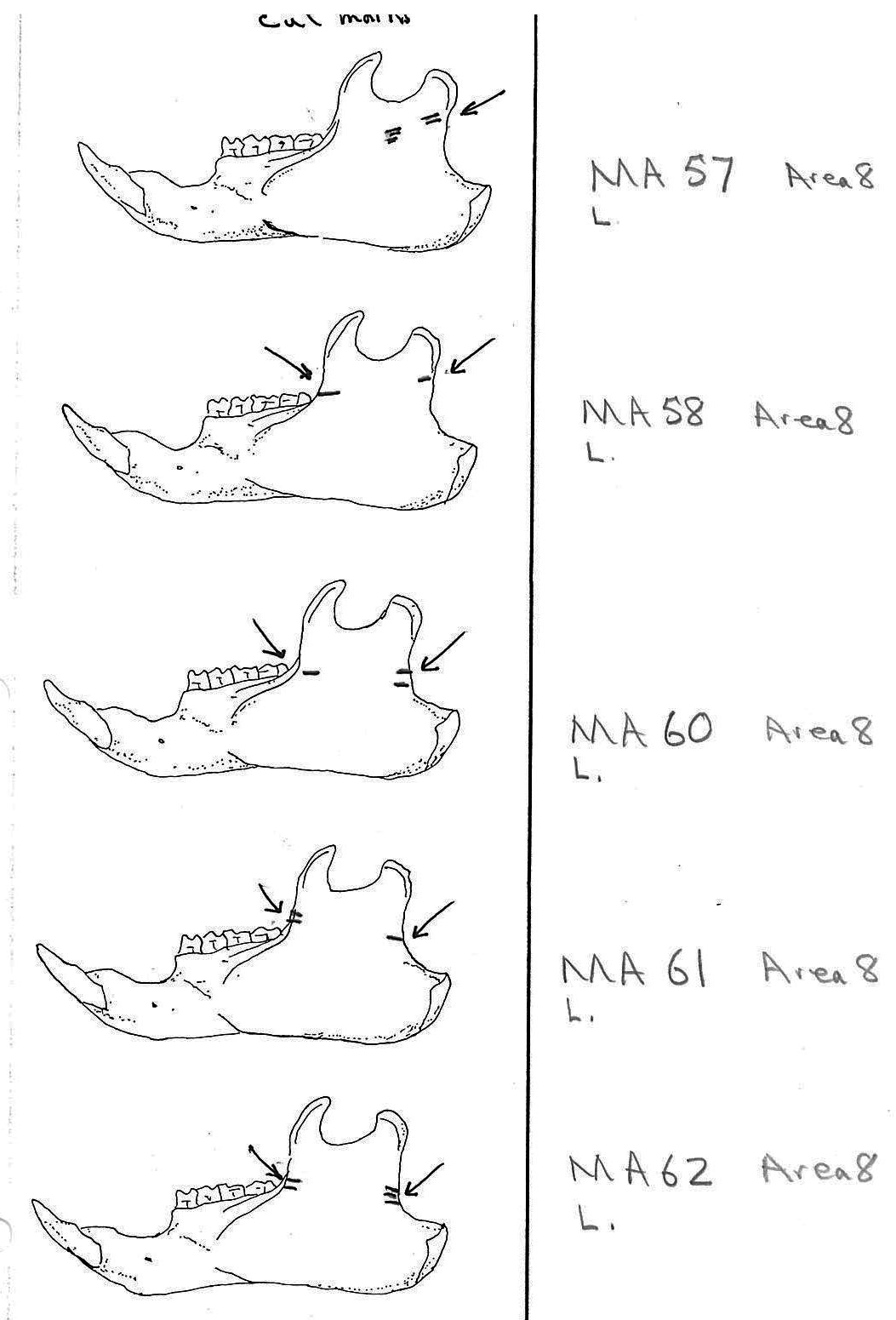
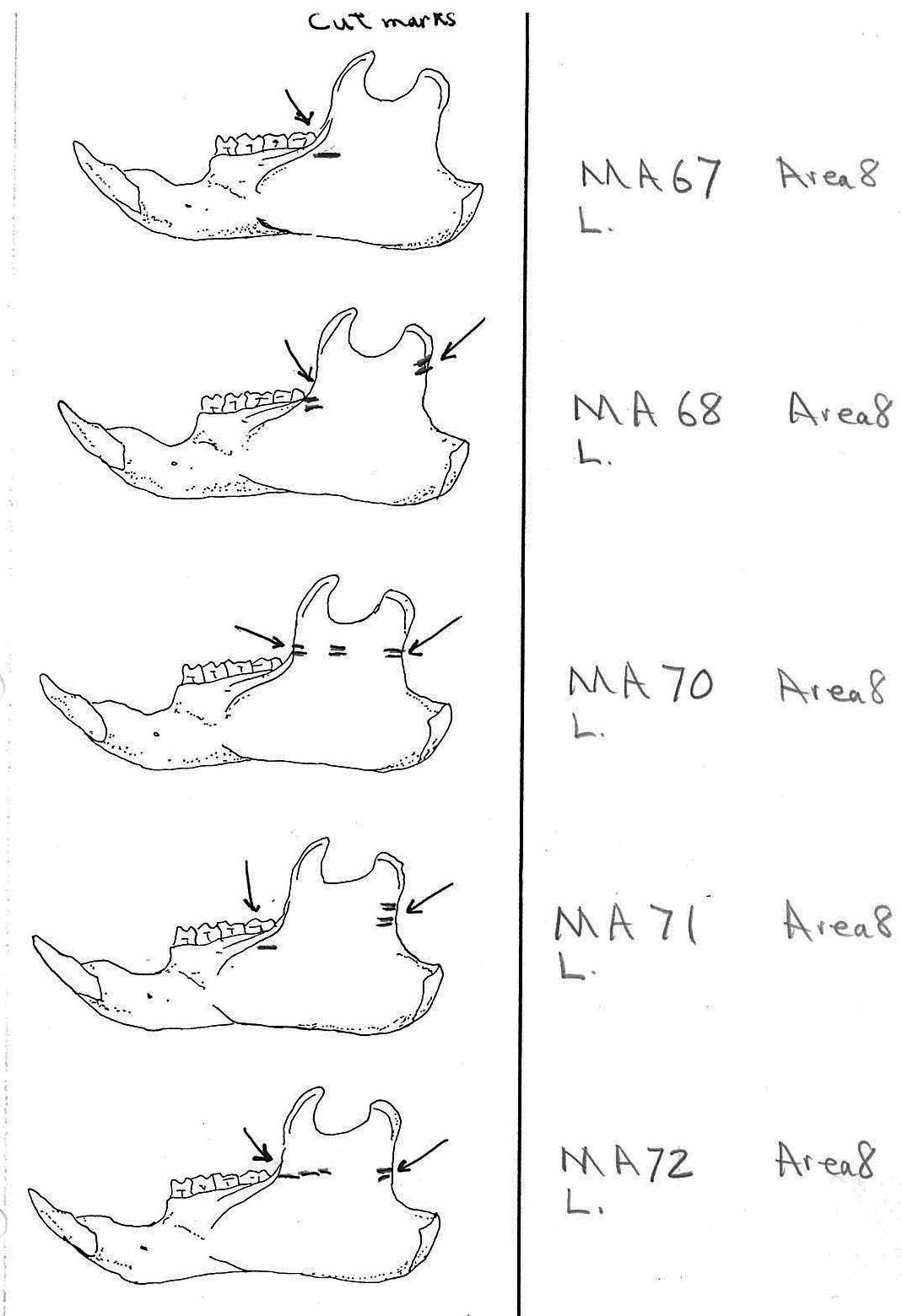
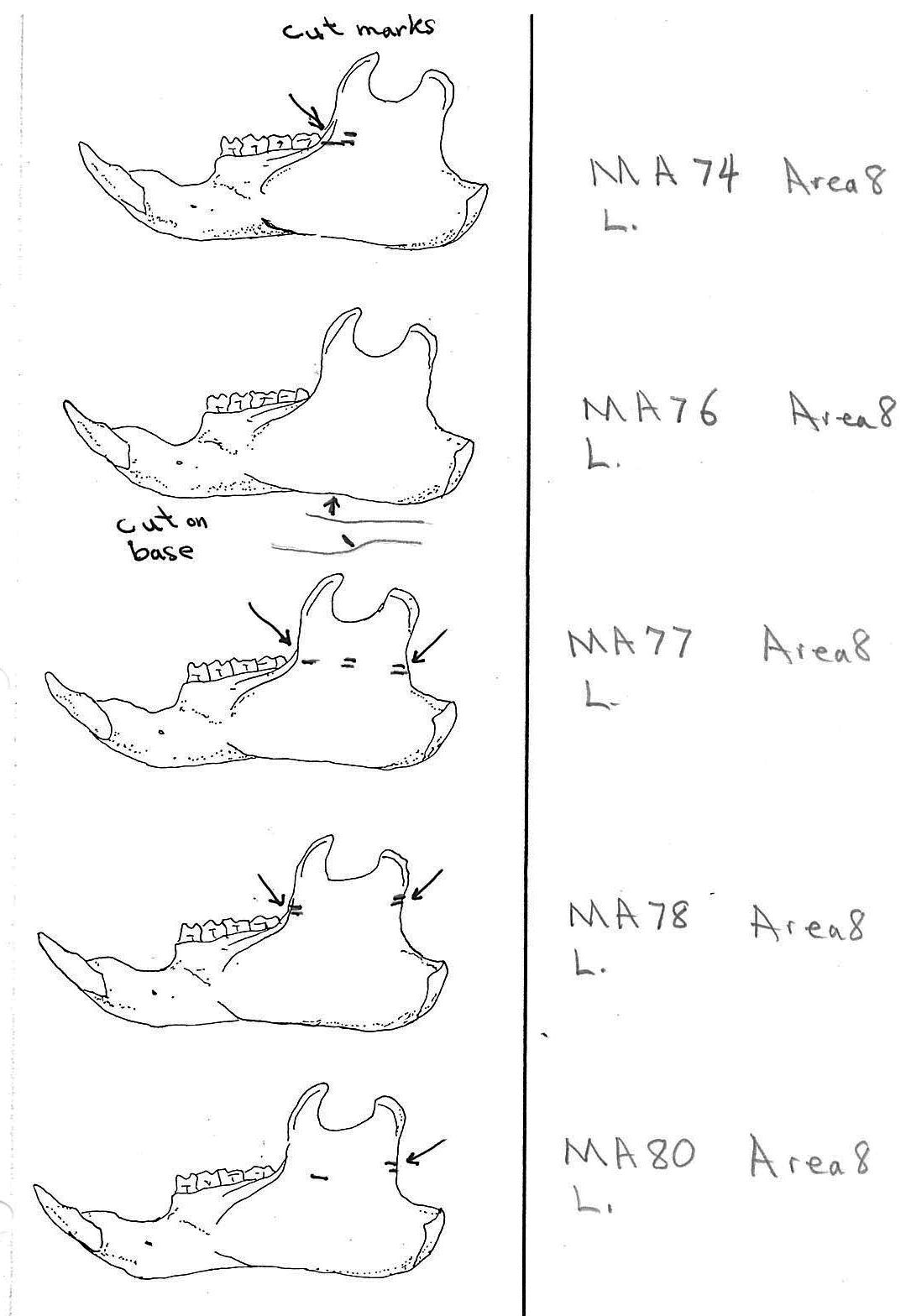
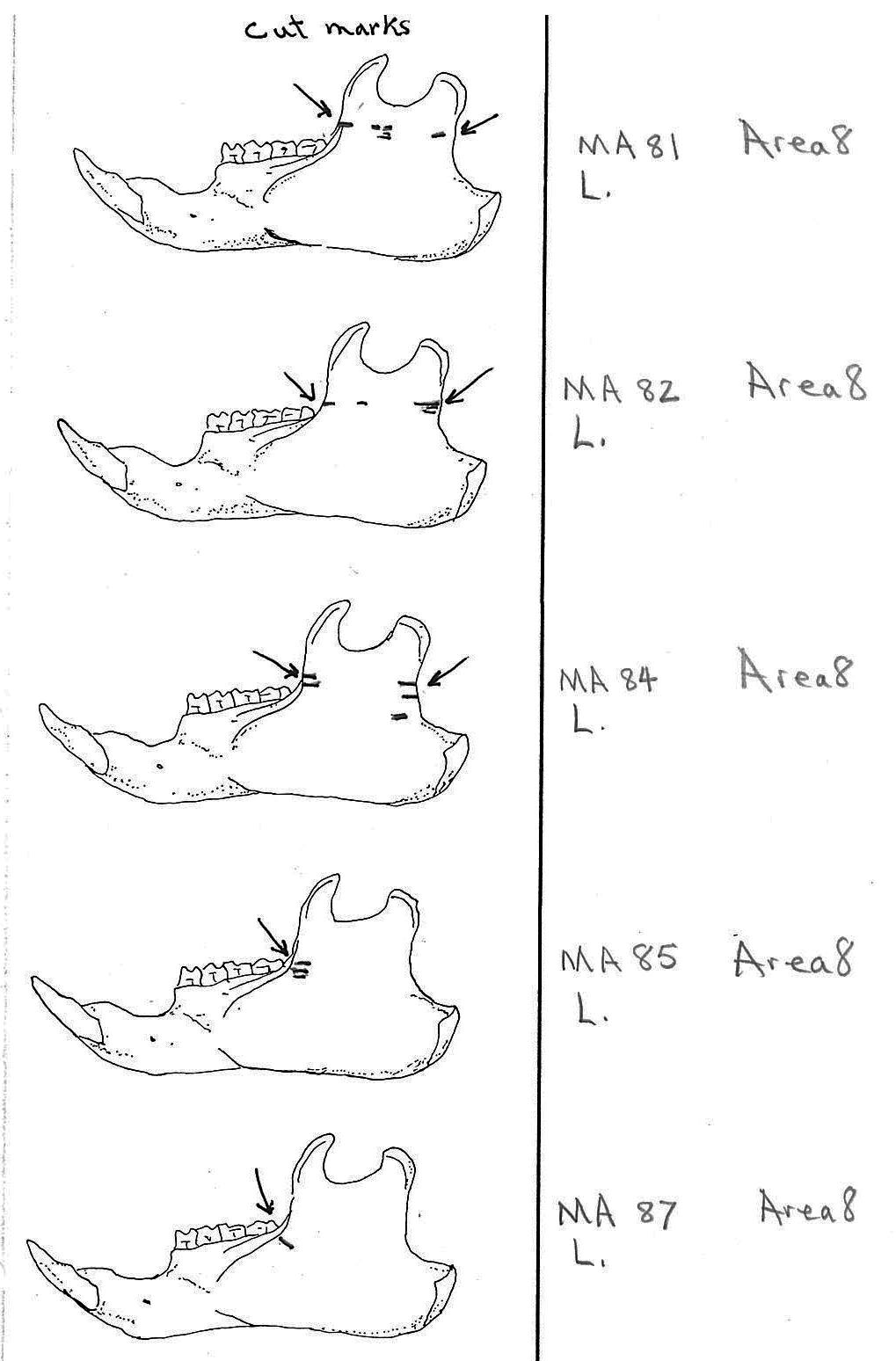
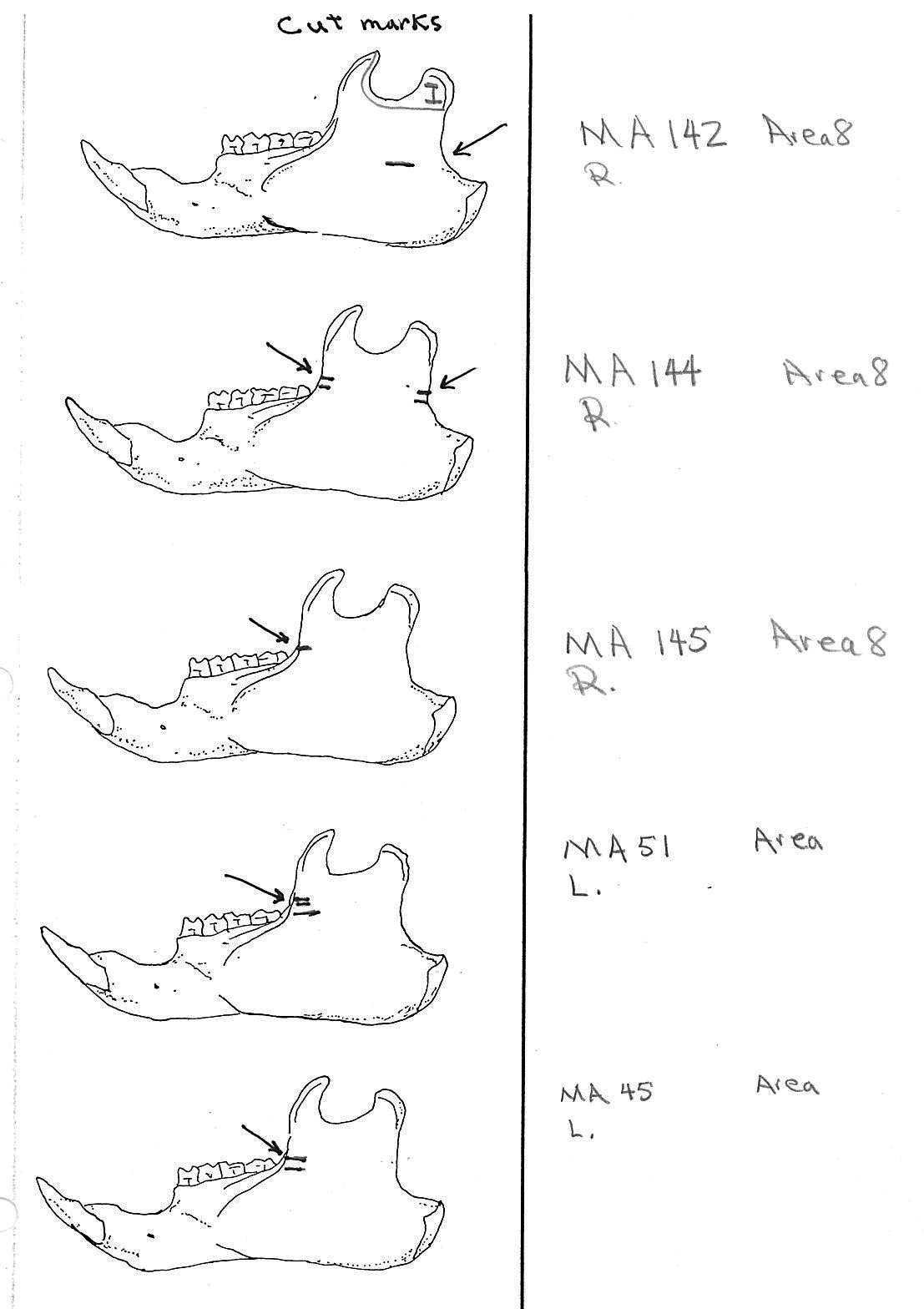
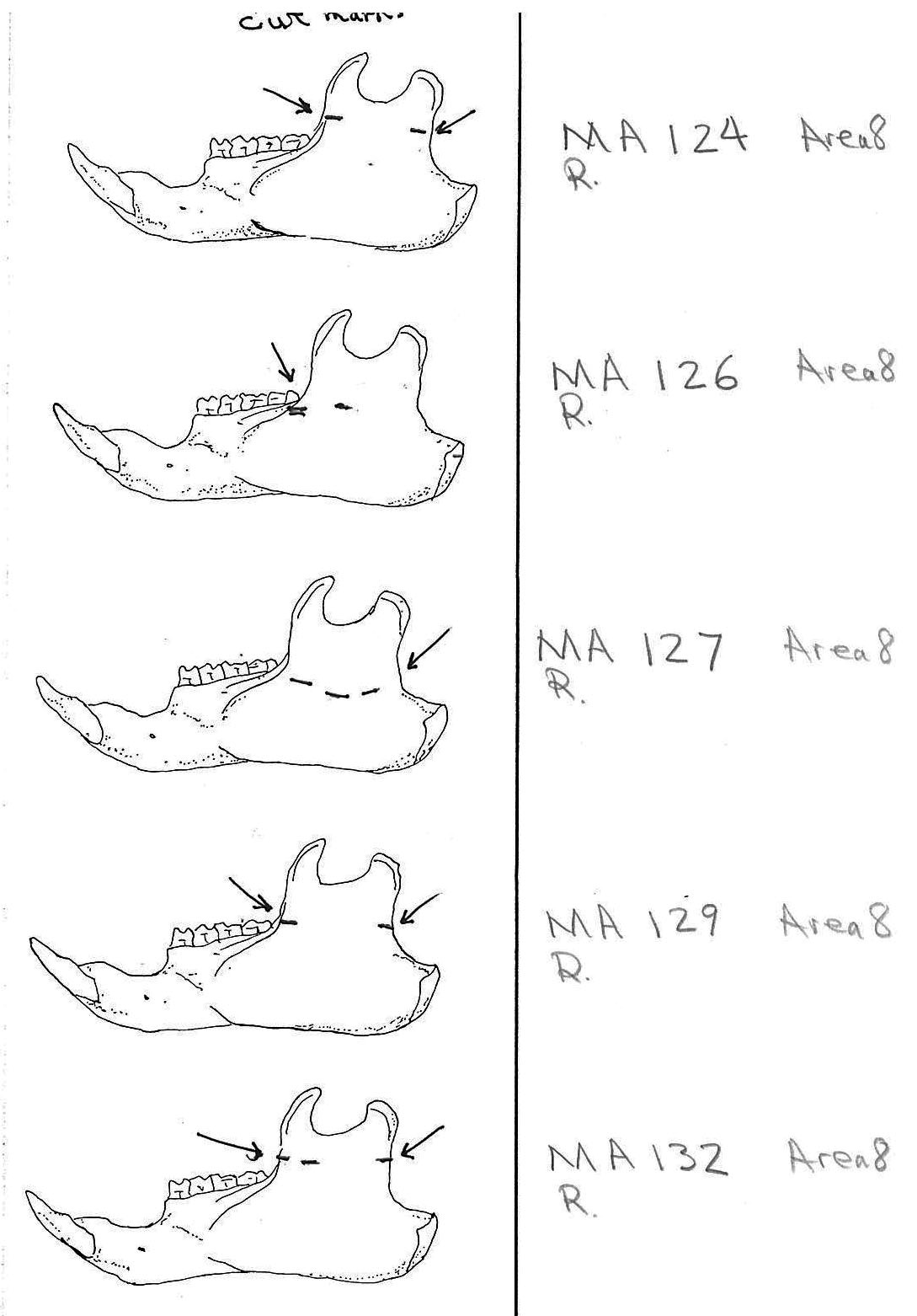
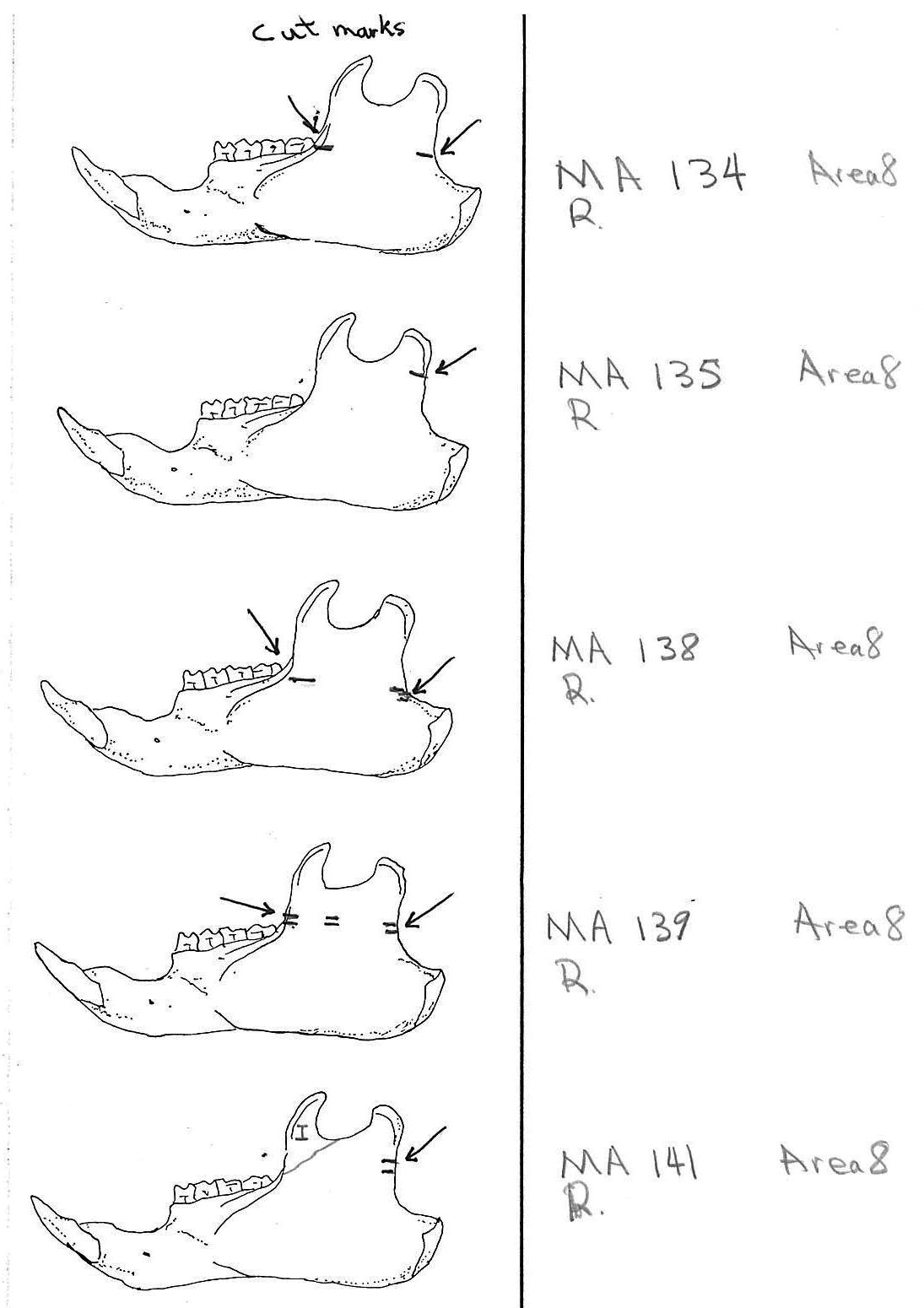
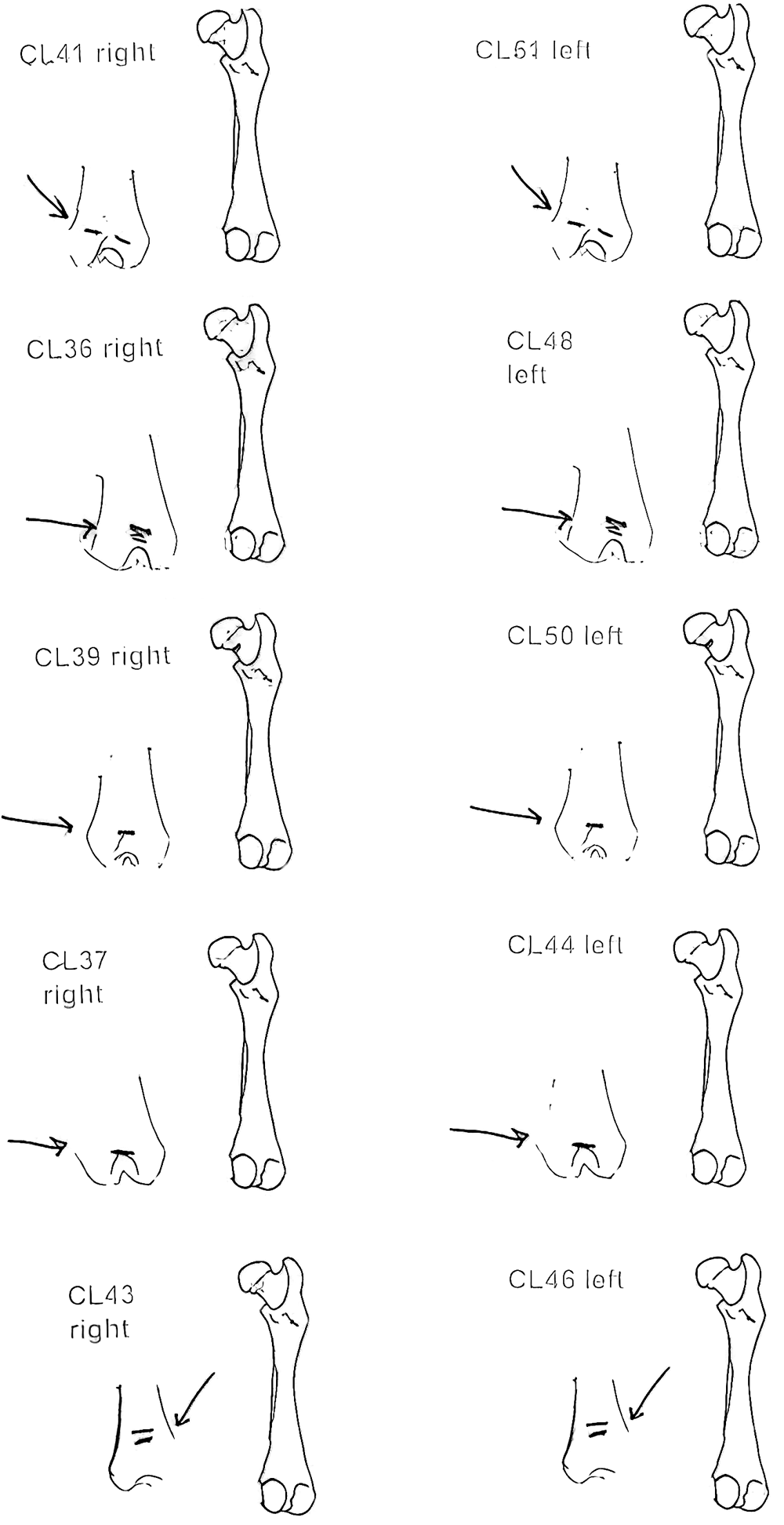
Modification Of The Femur
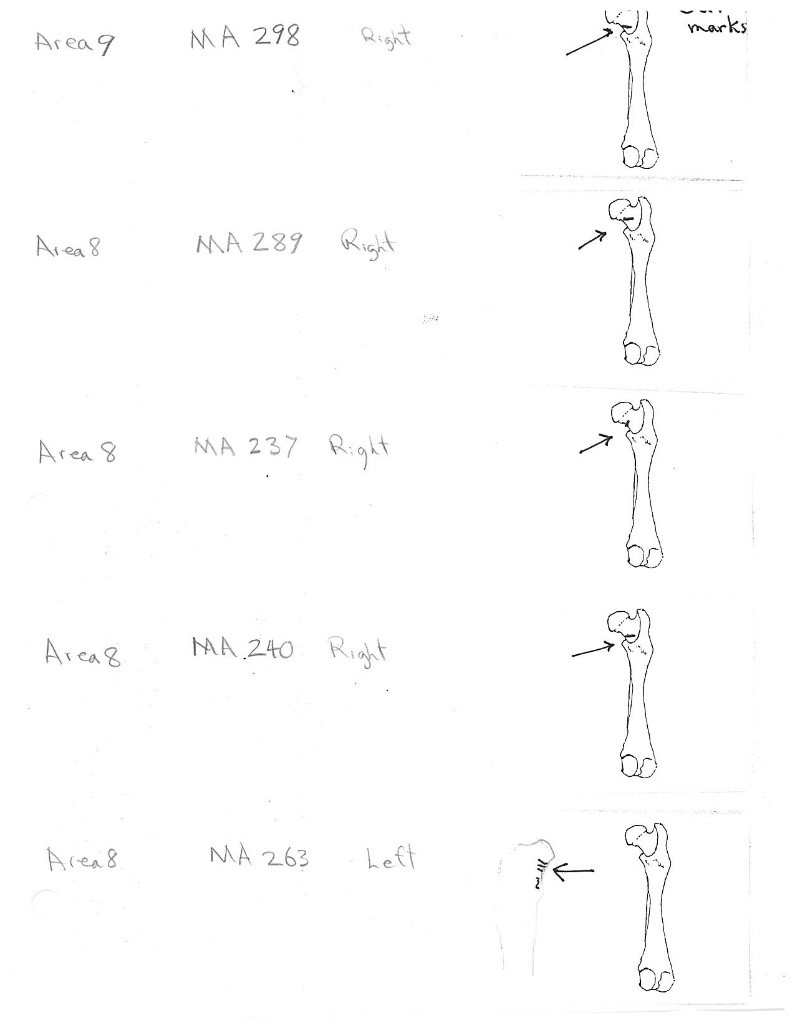
There are 22 femurs with cut marks. All but two of the bones have cuts that occur on the neck of the femur where the iliofemoral and ischiofemoral ligaments are located. Two of the later also have additional cuts. One has 3 cuts on the proximal shaft just at the base of the greater trochanter and another has a series of six cuts on the posterior side of the upper greater trochanter. One femur has only one cut across the front of the upper shaft and another has an upward cut on the outer trochanteric crest just above six small cuts at the top of the shaft. There are no cut marks on the distal end of femurs.

Modification Of The Humerus
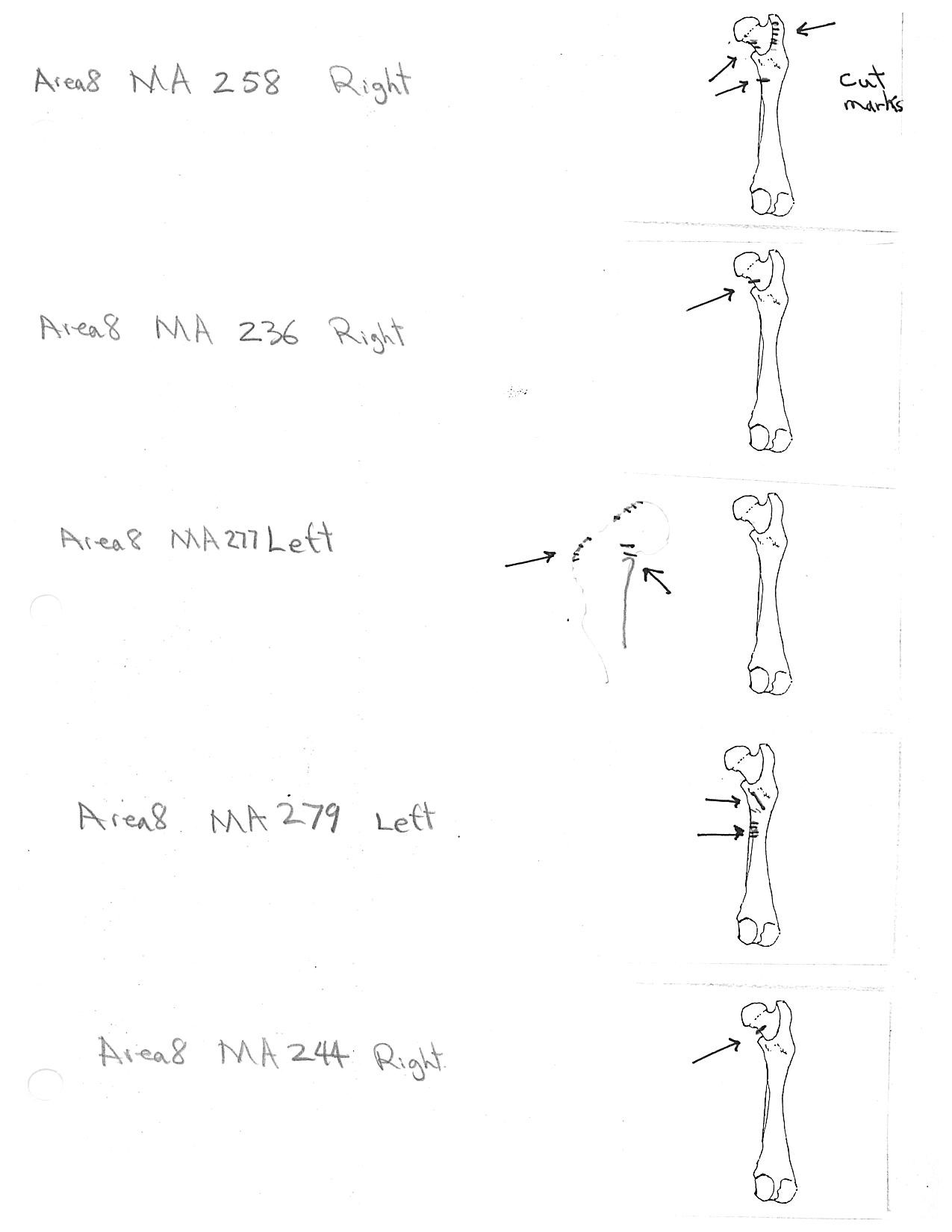
Four humerus bones have cut marks on the distal end. There are no cut marks on any proximal end. Three of the specimens, two left and one right, have one or two cuts across the small bone bridge above the lateral epicondyle on the anterior surface of the bone. One specimen has cut marks on both sides of the distal posterior side of the humerus. There are a few more cases of possible cuts and fractures on this muscle attachment bridge that have likely resulted from the pressure of a knife blade. In some specimens the small bone bridge is congenitally missing.
Modification Of The Tibia
There are six tibias with cut marks. Two have dubious cuts on the inside surface of the proximal shaft. Four have single cuts on the distal shaft, three on the posterior surface and one on the anterior surface. All of these cuts may be a result of skinning activities but their infrequency does not suggest any regular pattern.
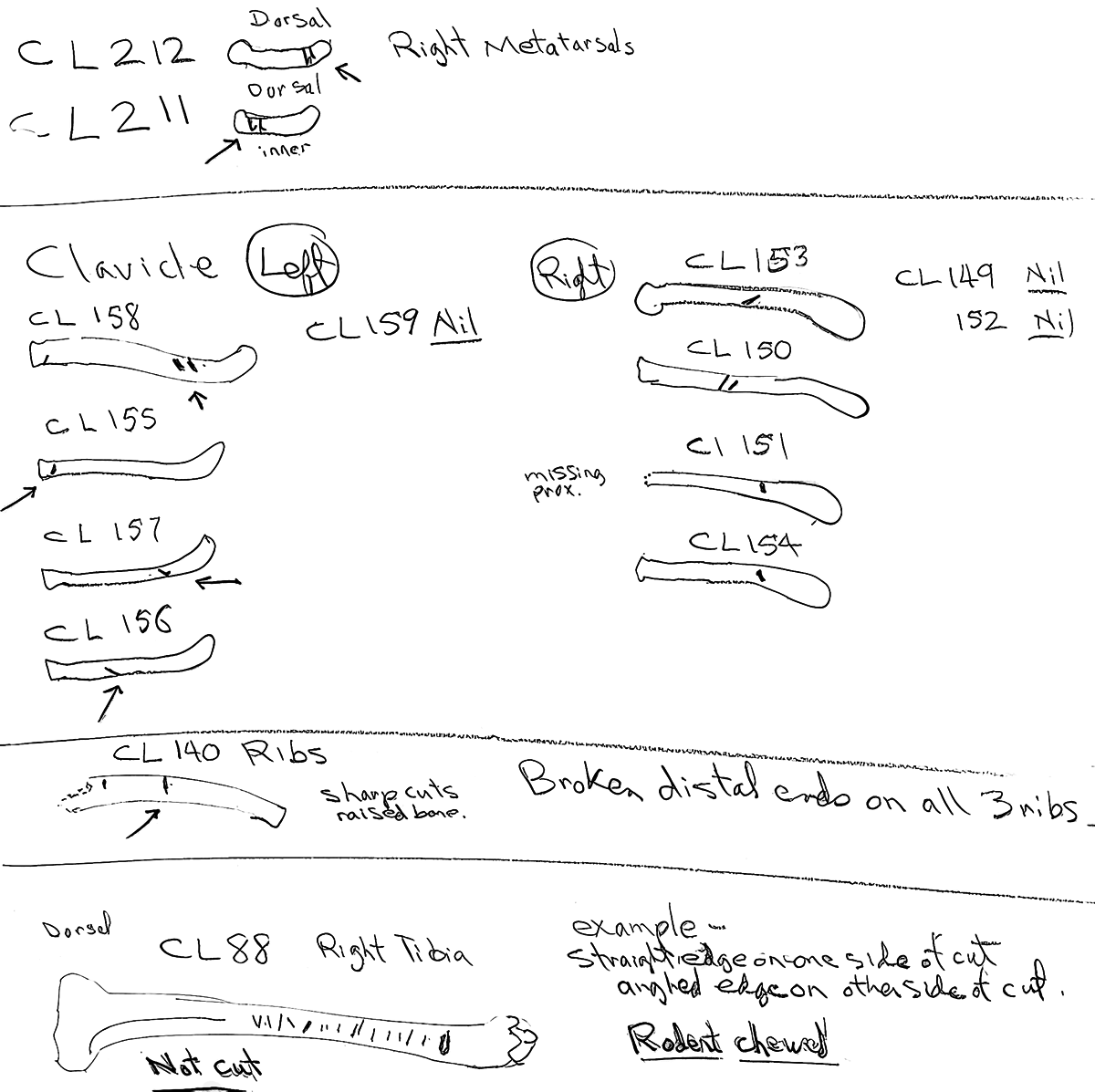
Modification Of The Ribs
Of a total of 168 ribs recovered three have single cut marks. Two are cut on the proximal end and one in the mid section.
Modification Of The Innominate
There is only one fortuitous cut on the inside surface of a right ischium. The innominate were removed without cutting the muscles attached to them. This would suggest than the abdominal area was likely snapped and pulled off the sacrum and discarded.
Martin, Deer and Bear Bone Cut Marks. Mariner Cave
Marmot Bones From Mariner Cave
SUMMARY LIST OF PRESENTS AND ABSENCE OF CUT MARKS
(MNI = Minimum number of individuals).
(Showing MNI represented by each cut element class and the total MNI in the site represented by that element)
| BONE ELEMENT: | NUMBER WITH CUT MARKS(S) |
|---|---|
| Mandible | 80 (MNI CUT 41) (site 74) |
| Cranium | 23 (MNI CUT 23) (site 62) |
| Femur | 22 (MNI CUT 14) (site 55) |
| Tibia | 6 (MNI CUT 4) (site 48) |
| Humerus | 4 (MNI CUT 2) (site 56) |
| Ribs | 3 (MNI CUT 1) (of 168 ribs) |
| Innominate | 1 (MNI CUT 1) (site 43) |
BONE ELEMENTS WITH NO CUTS: fibula; scapula; ulna; radius; clavicle; foot bones; vertebrae.
Additional qualifiers for cranium parts.
The total number of times (75) that cuts occur at a particular location on the individual craniums and non-matching parts of craniums is as follows: R. incisive 5; L. incisive 16; R. nasal 1; L. nasal 4; occipital condyle 15; R. bulla 16; L. bullae 18.
For comparative purposes – the cut locations on the 27 fragmentary cranial parts that do not count as individuals are located as follows: R. incisive 0; L. incisive 2; R. nasal 1; L. nasal 4; occipital condyle 8; R. bulla 6; L. bulla 9. Of the 75 cut location counts, 30 are on fragments (3 occur in combinations of two or more). Of the 23 cut cranium that count as individuals there are 45 counts (15 occur in combinations of two or more).
Missing Bones
Based on the presents of mandibles there were 74 marmots that were at least partially processed at the site location. The MNI represented by craniums suggests that 12 craniums are missing. These could be present in the assemblage but simply not showing up in the MNI count because of the fragmentary nature of some of the cranium. Two skulls, not collected, were later observed in rock crevices.
The larger long bones represent from 48 to 56 animals and the innominates only 43 individuals. This would suggest that the legs and pelvis were left on some of the animals attached to the main part of the carcass or removed and taken away separately. The numerous unfused innominate parts of younger animals would likely increase the MNI count to at least that of the average number of long bones.
None of the expected, close to 1500, coccygeal vertebrae or the sacrum are present. Of the expected close to 2000 vertebrae above the sacrum only a small number are present. Even given that many of the smaller vertebrae could have been missed in the collection process, the vast majority of the entire vertebral columns are missing from the site. Only a small number of the expected 962 ribs are present. This evidence clearly shows that the backbone and rib cage were consistently separated from the carcase and removed from the site.
Only a few of the approximately 3000 expected foot and paw bones are present. Although some of these very small bones were likely missed in the collection procedure, most are missing and assumed to have been left in the pelt during the skinning process.
Distribution Of Marmot Bones By Collection Areas Of Cave
The way marmot remains were lumped together in nine areas of the cave gives some clues to the nature of the marmot processing activities. The majority of bone elements were located in Area 8.
Area 1 (MNI 10). This appears to be an area where the complete skull (cranium with mandibles) of about ten marmots were removed at the same time and dumped. Ten well preserved skulls and fragments with 8 right and 8 left mandibles were found close together. The post cranial bones in Area 1 represent only small portions of a maximum of four individuals based on humerus, tibia and femur elements, and fewer animals based on all other bone elements. Six innominates are included in the latter.
Area 2 (MNI 3). There are no skull parts or femurs in this area. A minimum of three marmots are indicated by three different sizes of scapula in the 20 postcranial bones.
Area 3 (MNI 10). This area includes ten right and eight left mandibles with a minimum of four animals based on skull sections. Other remains include 9 long bones and four innominates from three individuals based on right humerus. Area 3 is similar to Area 1 in having mostly skulls parts and some long bones.
Area 4 (MNI 9). This area has 9 individuals based on cranium and fragmentary non-matching cranial parts but contains only one mandible. Postcranial bones include a significant amount of 20 ribs along with 4 right humeri, 6 innominates and little else.
Area 5 (MNI 5). Five cranium and 6 left and 3 right mandibles are found in this cluster. Others include about 30 long bones and, significantly, about 30 ribs.
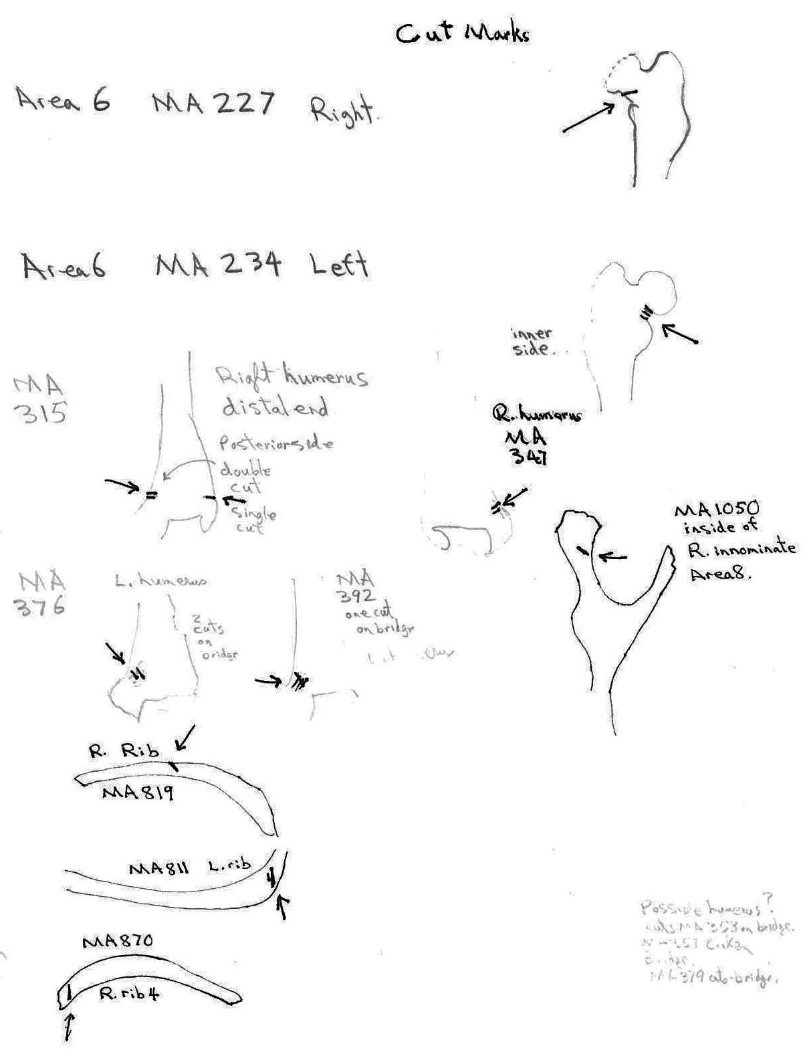
Area 6 (MNI 7). This area includes mainly long bone portions (44) of seven individuals. Only 3 cranium and two sets of mandibles are present. Six innominates are present.
Area 7 (MNI 3). This small pile includes 2 cranium and three mandibles, 2 left humerus with no epiphysis and 1 left with epiphysis, and a matching left and right innominate.
Area 8 (MNI 44). The vast majority of bones occur in this large cluster. Skulls are less frequent in this area in proportion to post cranial remains than in the other areas.
Area 9 (MNI 2). This is an area of 21 loosely scattered bones that did not form as distinct a cluster as the other area samples. It included 2 left and 2 right tibia and the same for mandibles.
The craniums in all collection areas except area 8 may represent as many as 33 individuals, but it is uncertain how much overlap there is between clusters. Even given some overlap there is still a minimum of about 25 craniums that would be exclusive of area 8. This clearly shows that a large number of the 62 craniums from the site are located in six of the smaller areas of bone concentrations. If each of the nine Areas were given a separate MNI count it would add up to 93 with 44 in Area 8. This figure would also tend to suggest that craniums are underrepresented in Area 8.
Conclusions Deduced From The Distribution, Absents And Modifications Of Marmot Bones
Skinning marks indicate that the fur was separated from at least the head and legs. The front legs appear to have been cut into, at least part of the time, near the distal end of the humerus. The lack of cuts on the ulna and radius suggest they were pulled out to leave the carpals, metacarpals and phalanges attached to the fur. The back legs appear to have been occasionally cut into near the base of the tibia and pulled separate from the foot bones which were left attached to the fur.
Butcher marks indicate that the tongue was cut from the head, the masseter muscles cut to separate the jaw from the cranium or to leave the larger part of the muscle attached to the neck for its food value. The front legs, scapula and clavicle were separated as a unit by cutting the muscles across the clavicle.
The back legs were separated from the pelvis by cutting the muscles at the femur neck. Since the femurs were discarded it would appear that they were removed to make the separation of the pelt easier.
The absence of cut marks on the pelvis would suggest that the ventral part of the abdomen was cut and the abdominal area snapped back and pulled off the sacrum. It can not be determined if the muscles attached to the sacrum were cut as the sacrums are missing.
The missing tail bones would suggest that the tail was left attached to the pelt or cut off and taken away. The vertebrae and ribs are almost all missing and therefore assumed to have been left intact. In a few cases the rib cage was removed and left behind.
This evidence would suggest that the animals were either: (1) entirely skinned and then butchered in such a way as to leave the bulk of the fat and meat in one piece attached to the backbone and ribs. These products were then taken to another location. (2) Partially skinned and partially butchered. The partially separated loose fur may have been pulled over the ends and tied across the intact fur still attached to the backbone with ribs.
The uneven distribution of body parts in the different cave clusters may suggest that different individuals processed different parts of the animals. Some of the smaller clusters which contain many skulls and some leg bones may have been created when one person undertook the job of removing the fur from the skull and then removed the skull and legs before passing the remains on for someone else to finish the processing.
The uneven or fragmentary distribution of postcranial elements in the smaller bone clusters suggests that parts of the same animal were thrown on different piles during the butchering process and that the distinct bone clusters do not necessarily represent different time periods or distinct episodes of animal processing.
Area 1 clearly appears to represent a portion of one marmot processing event where ten skulls were cut off and placed close together on the same level in a small concentration. When one looks at the positioning of the bone clusters in the cave and their contents, there are two zones of three bone clusters each which represent about 10 animals each. These include the smaller adjacent clusters 2, 3 and 4 as one potential processing zone and clusters 5, 6, and 7 as another zone. The killing of about ten marmots at a time would suggest that the bones represent about 7 hunting episodes at this location. This would fit the norm for the number of animals in a typical colony.
Modifications To Bear Bones From Mariner
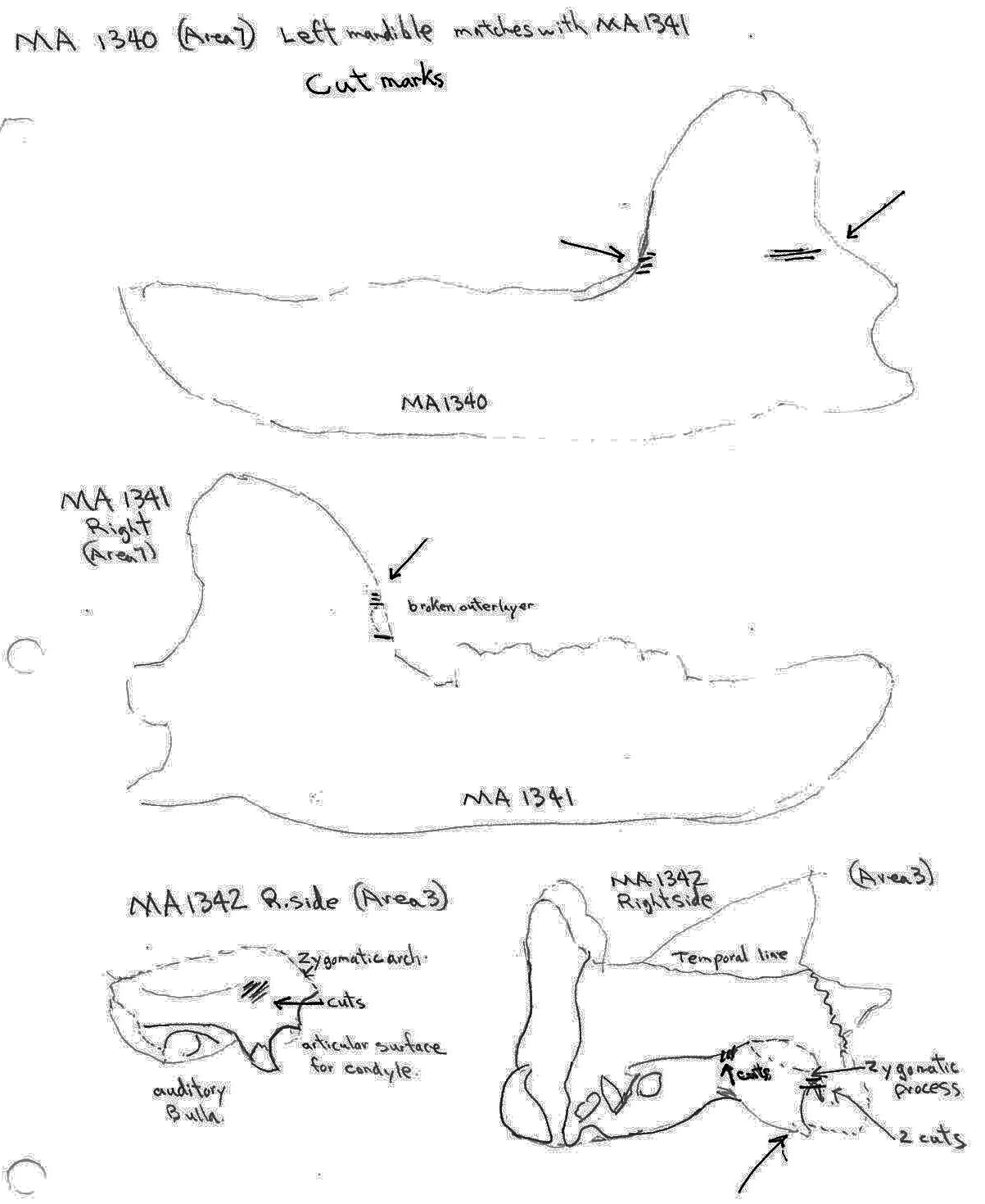
There are cut marks on three bear skull fragments. Two right posterior skull fragments (MA1342a and MA1342b) have cuts across the posterior end of the zygomatic process and one left maxilla fragment (MA1358) has an upward cut forward of the infraorbital foramen.
Five bear mandibles have cuts on one or both sides across the base of the ascending ramus. Two mandibles, MA1340 and MA1341, are a matching pair. The left one shows cuts on the posterior and anterior surface of the ascending ramus while the right has cuts marks only at the anterior side.
Only two post cranial bones show evidence of human modification. A left scapula has three cut marks on one edge and a right radius has a series of very fine cut marks just below the distal end. Gnaw and tooth pressure marks are also found on the latter bone.
Modifications To Deer Bones From Mariner
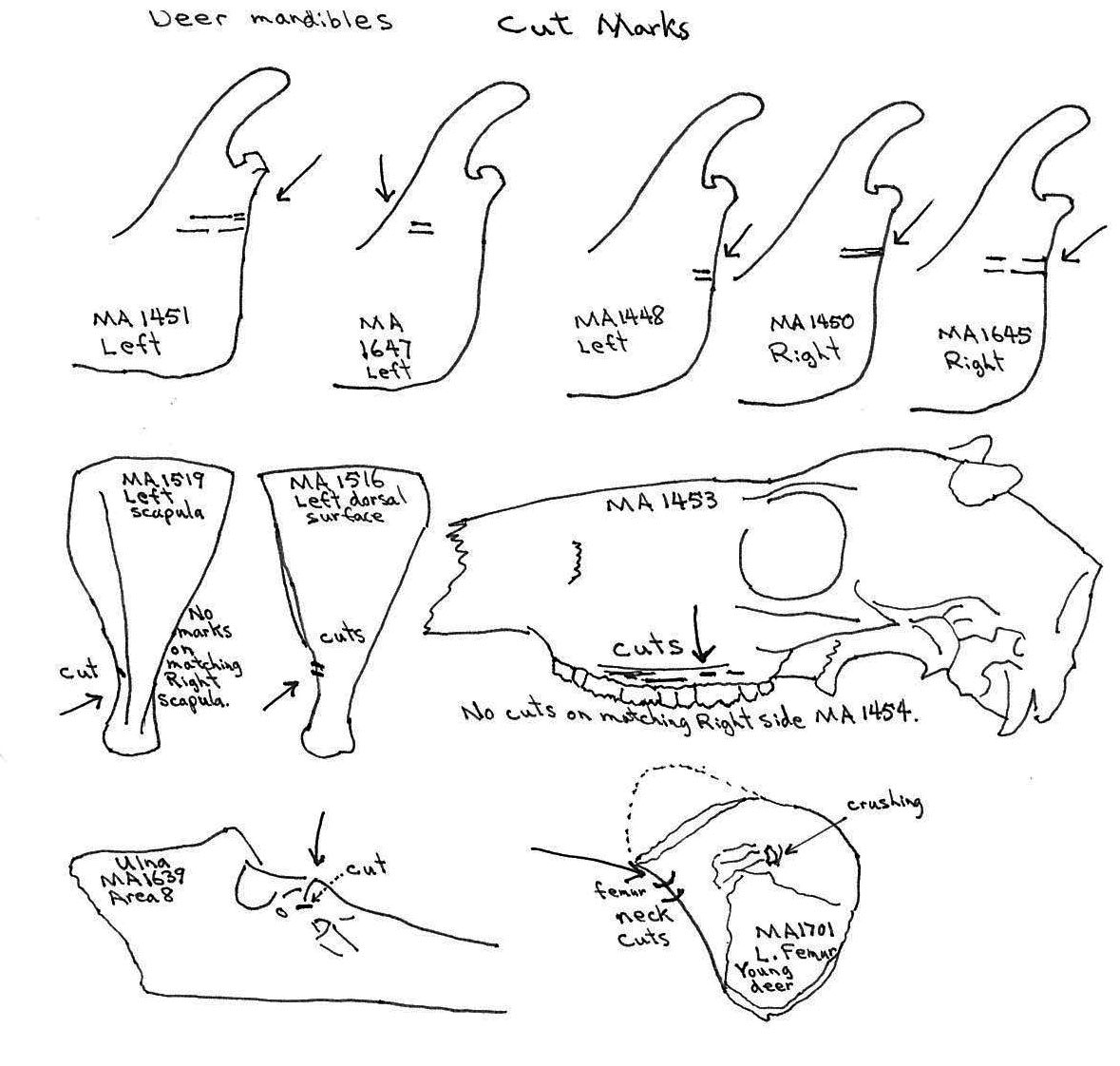
A deer skull (MA1453) has a series of parallel skinning cut marks on the left side above the tooth row on the maxillae. No cuts are found on the corresponding right side. Five deer mandibles have cut marks across the posterior base of the ascending ramus.
Of 7 right and 5 left scapulas, cut marks are found only on two left scapula – one on the outer (MA1516) and one on the inner edge (MA1519) of the constricted area below the articular end. Two cuts are located on the neck of a left femur MA1701 but none are found on the other five proximal ends, the four distal ends or the many shaft fragments.
Most ulnas are not cut, only a single cut can be observed between the articular surfaces of MA1638. Of four humerus fragments a cut is seen on the distal shaft of a humerus MA1437, on the distal left humerus MA1703 at the junction of the shaft and two cuts on the inside of the greater tubercle of a right humerus MA1349.
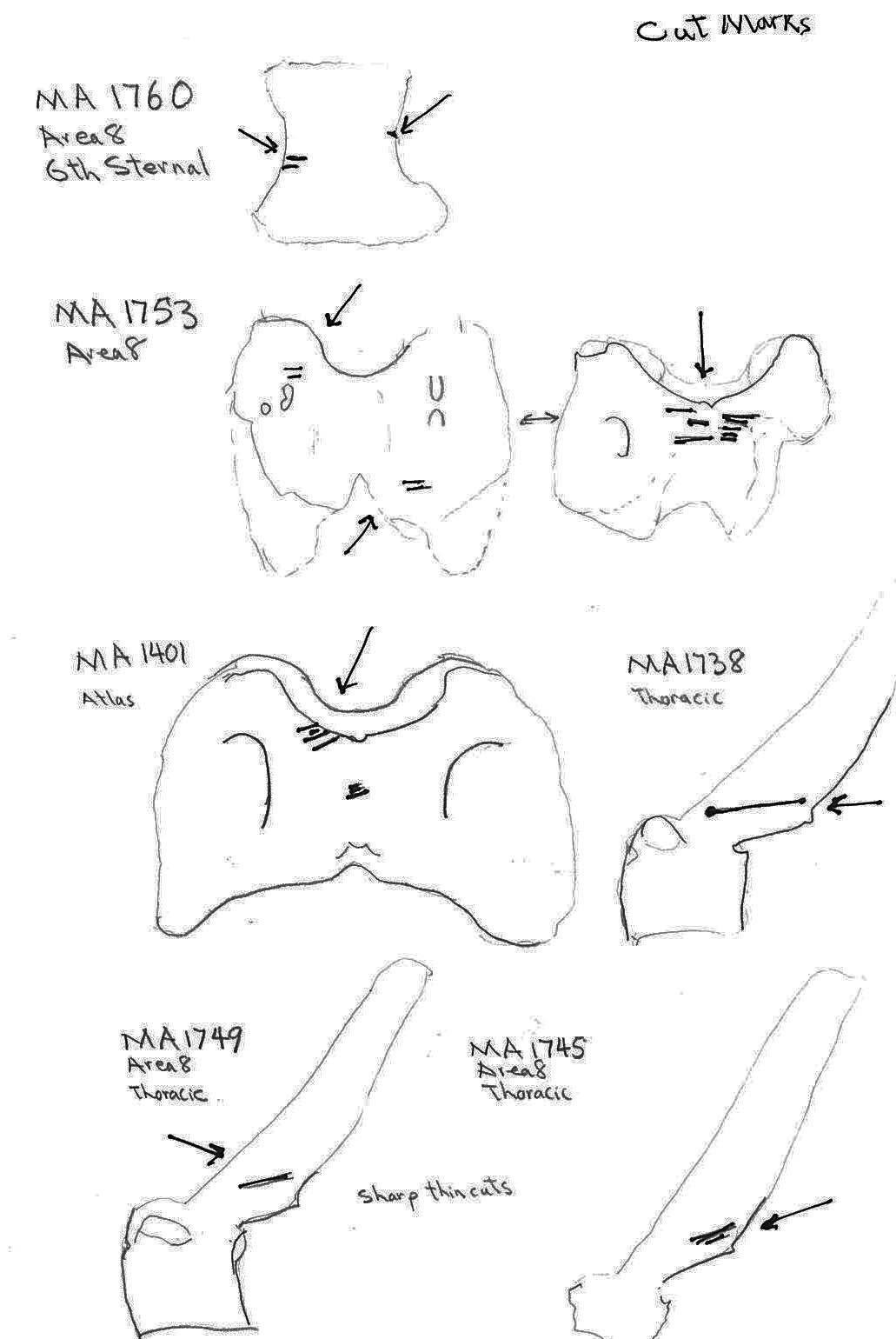
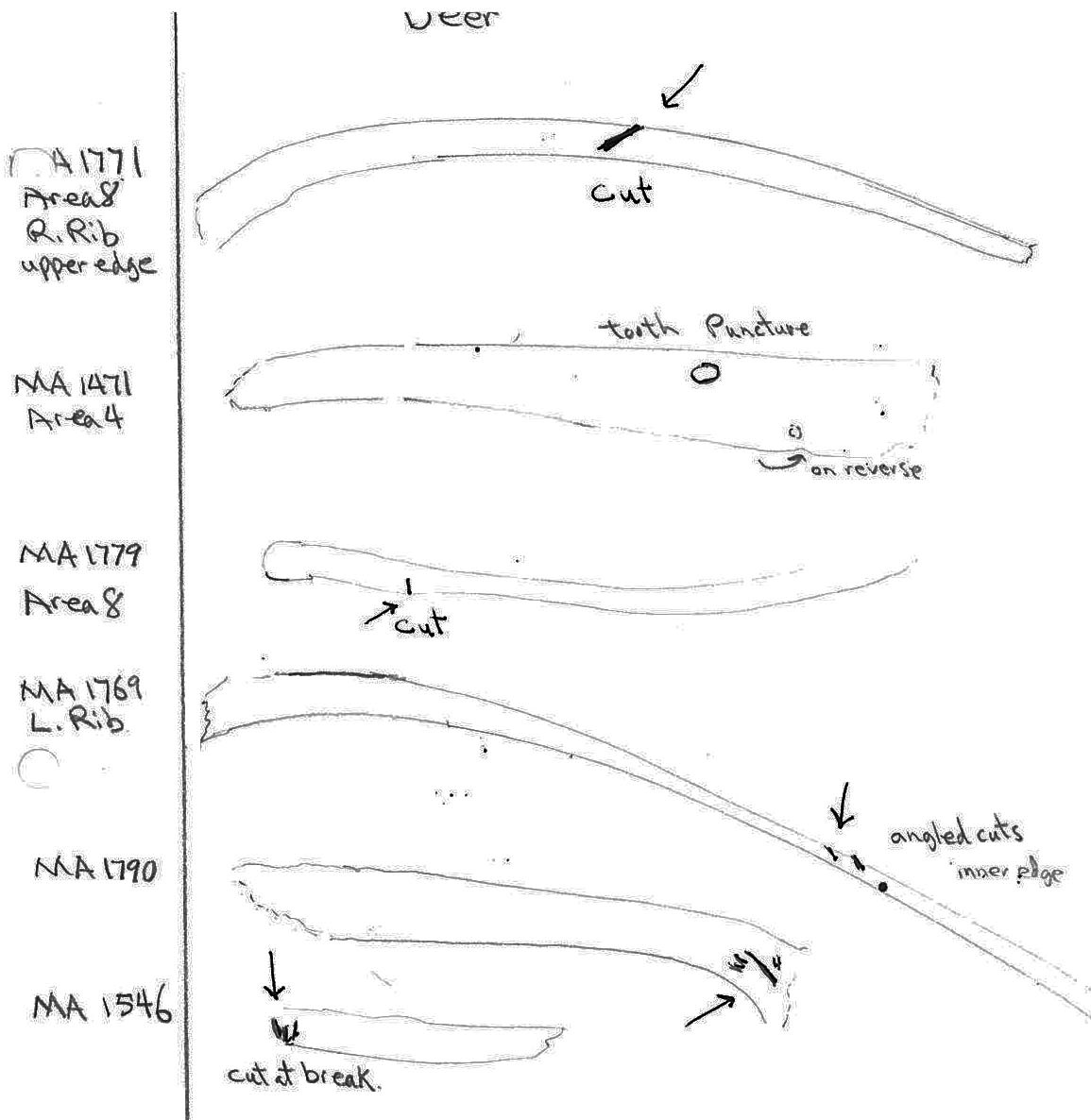
Five ribs have cut marks. Two (MA1771 and MA 1769) are cut across the middle of the ribs; MA1790 and MA1779 are cut near the proximal ends and MA1546 is a fragment cut at a broken edge. Six vertebrae have butchering cuts. The three thoracic MA1738, MA1745 and MA1749 are cut across the base of the spine; a 6th sternal (MA1760) is cut on two edges of the centrum; and two atlas vertebrae (MA1401 and MA1753) have multiple cuts across the upper ventral edge and with the latter also across the upper and lower dorsal edge. No cut marks (MNI 5) were observed on innominates.
Cut and Carnivore Chewed Deer and Bear Bones
Modification To Martin, Squirrel And Blue Grouse Bones
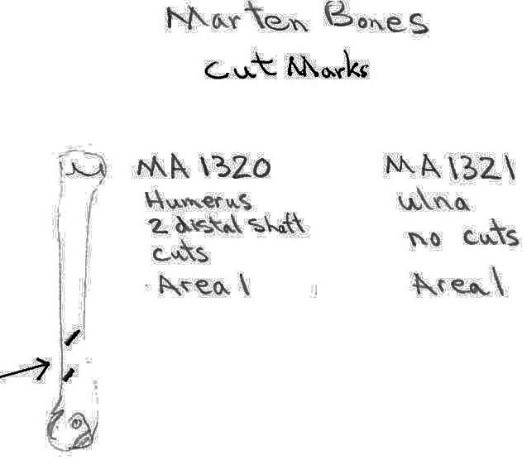
Two marten bones include an ulna with no cut marks and a humerus with two cuts on the distal shaft. The squirrel humerus and the blue grouse breast bone had no cut marks.
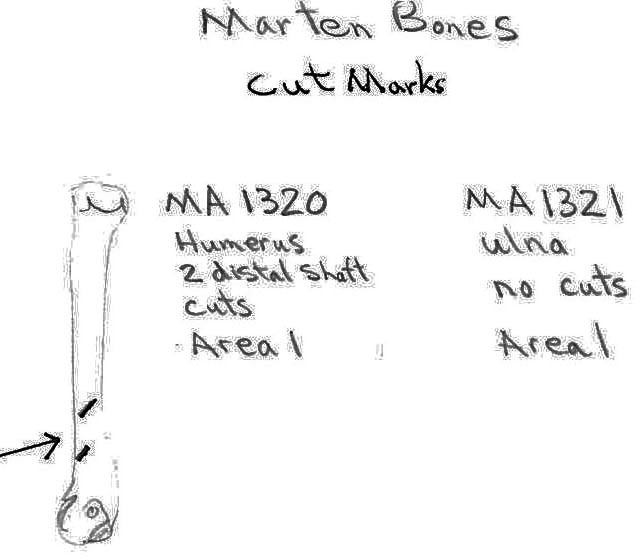
Modifications To Non-marmot Bones – Conclusions
Bear, deer and marten were all at least partially skinned and butchered in Mariner cave. The small sample size and sporadic nature of most of the cut marks does not allow us to observe sufficient evidence for any standard skinning and butchering procedure as is the case with the marmot bones. The skulls of bear and deer were skinned, removed and left at the site. Most of the post cranial remains, however, appear to have been cut up and removed from the site. It is conceivable that some of the bones were animals or parts dragged into the cave by carnivores but the presents of both carnivore chewing and butchering on a few bones would suggest that parts of these larger animals were either given to dogs or scavenged by carnivores after being abandoned by the hunters.
Tools Associated With The Mariner Cave Bones
Four artifacts were found in the Mariner Cave site. A sharpening or abrading stone made of coarse sandstone was found in the middle of Area 4 in a pile of marmot bones. This 11.5cm long rectangular stone tappers from 3.5 cm in width (1.5cm from one end) to 2.6cm at the other end. Its thickness varies from 1.37cm to 1.15cm at the narrowest end. The stone has a quartz groundmass with magnetite and mica present. It is ground on both flat sides but is extremely smoothly ground on one side. The rougher surface has black organic material adhering to it in parts. This tool could have been used to re-sharpen a ground mussel or slate knife. It is too smooth to have been used for primary sharpening of stone, bone or wooden projectile points.
A partially burnt knot of wood, possibly used as a torch, and chunks of charcoal from it, were found on top of the Area 5 bones. A deer scapula, burnt and heavily covered in charcoal soot, lay immediately below it.
Fragments of a California mussel shell were found together in Area 8, against the back of the west wall, in a pile of marmot bones. This is likely a part of a shell knife but does not include the distal end which usually forms a sharp cutting edge. This tool could have been used in the skinning process.
A green siltstone flake (L.4.42cm x 3.54cm x 1.55cm), with several possible working edges, was found in a pile of marmot bones in Area 8. This may have been used as a cutting tool for skinning and butchering. The flake has one long straight distal edge (2.85cm long) that continues in a curve to one side. This edge has slight rounding that may be natural weathering or use polish. There is no retouching on this edge. One side has step fracturing from earlier removal of smaller flakes as well as possible retouch at the bottom edge. Another short straight edge is slightly rounded with no obvious retouch. Two other short sections of the flake have what appears to be unifacial use retouch. The finest edge of this flake can be seen to fit the larger cuts on some bones, when both are observed together under a microscope. Other bone cuts are much finer and could not have been cut with this tool.
The width of some cuts is too fine for even the width of the edge of a microblade. This suggests the possible use of a fine ground iron blade for butchering. When fine metal blades cut into bone from the side, or obliquely, they often leave an overlapping small shelf of bone that remains in place. Cuts of this nature can be observed on some of the Mariner cave bones.
Human Alteration Of Bones From The Limestone Rock Shelter
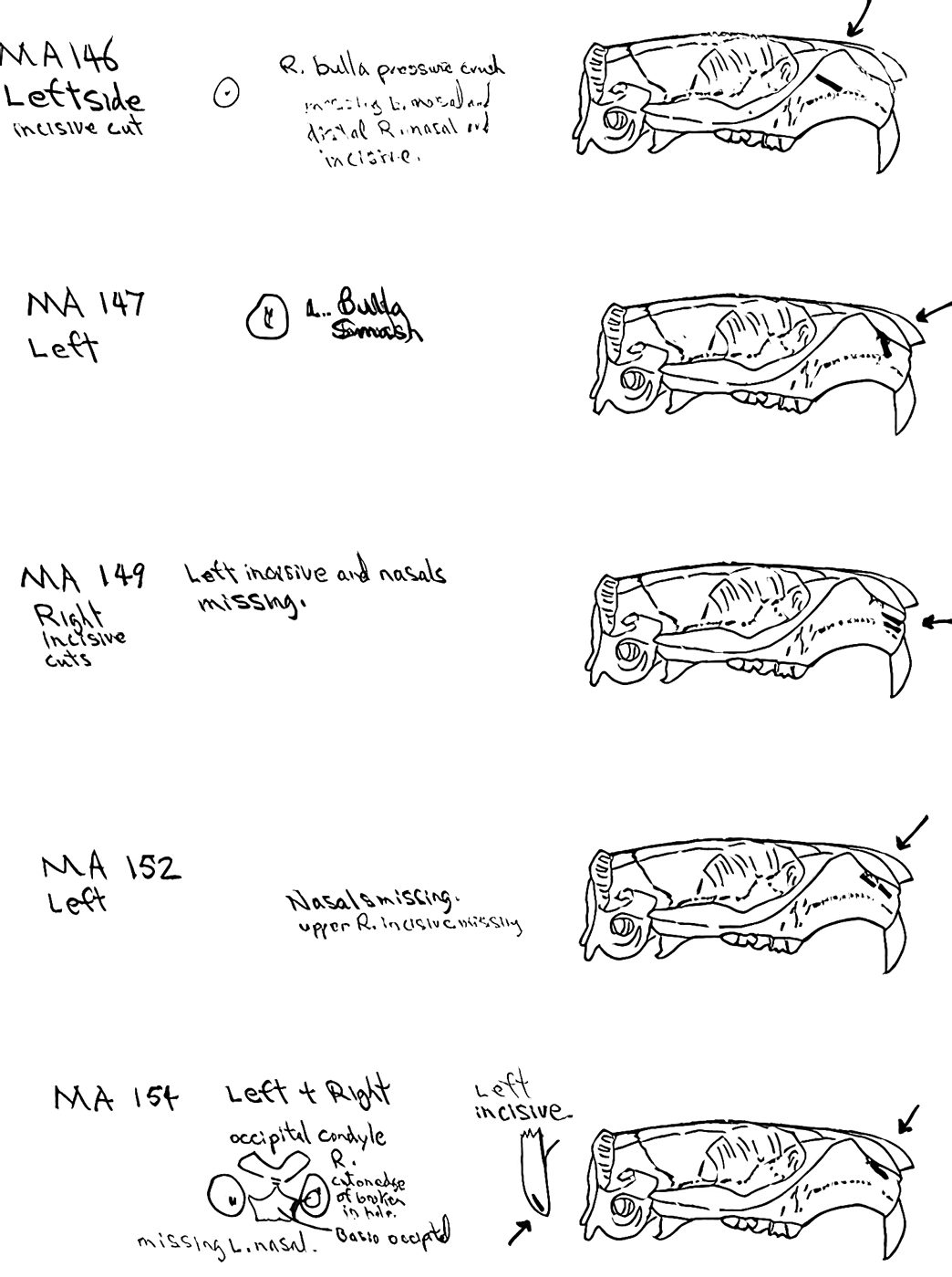
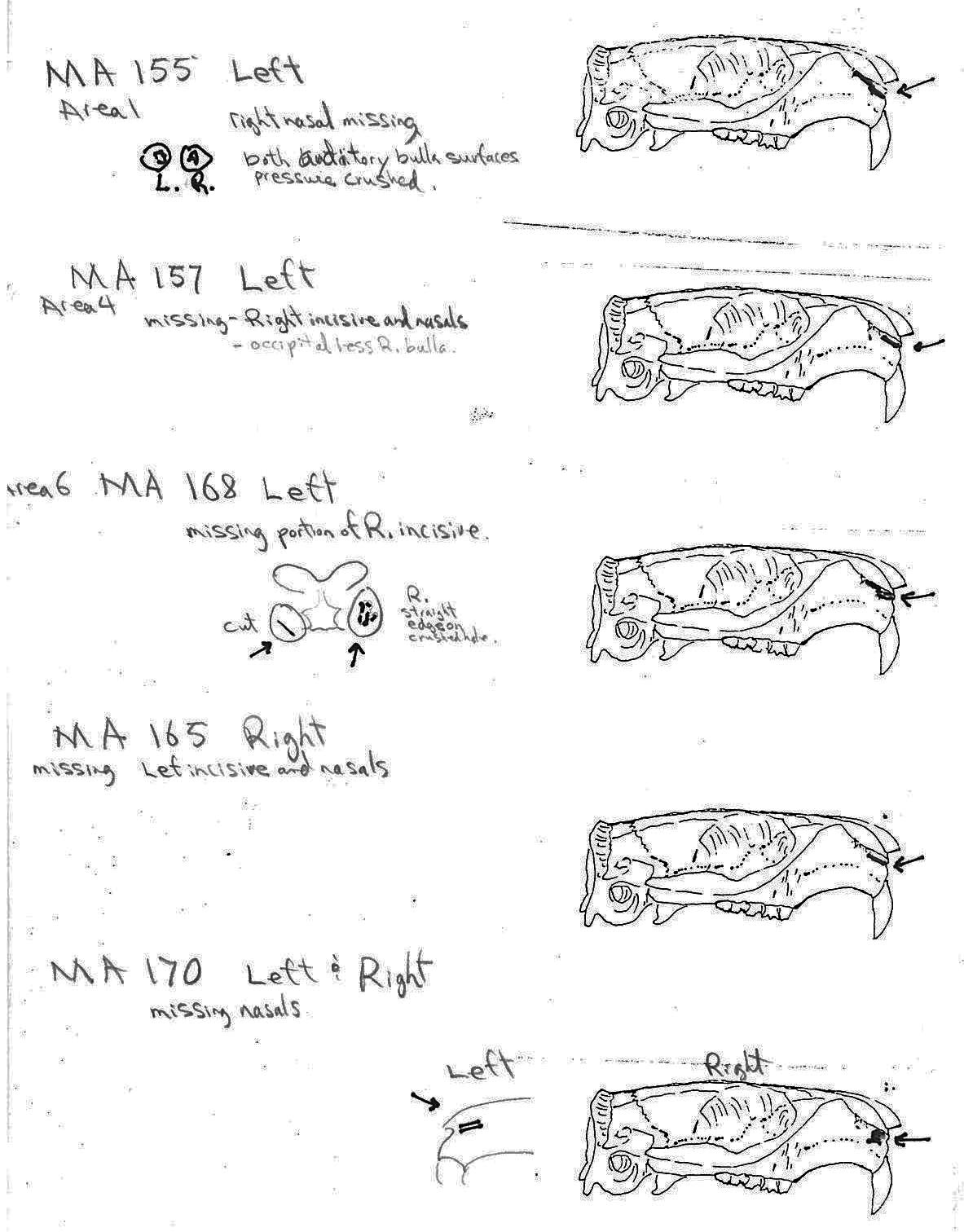
Modification Of The Cranium
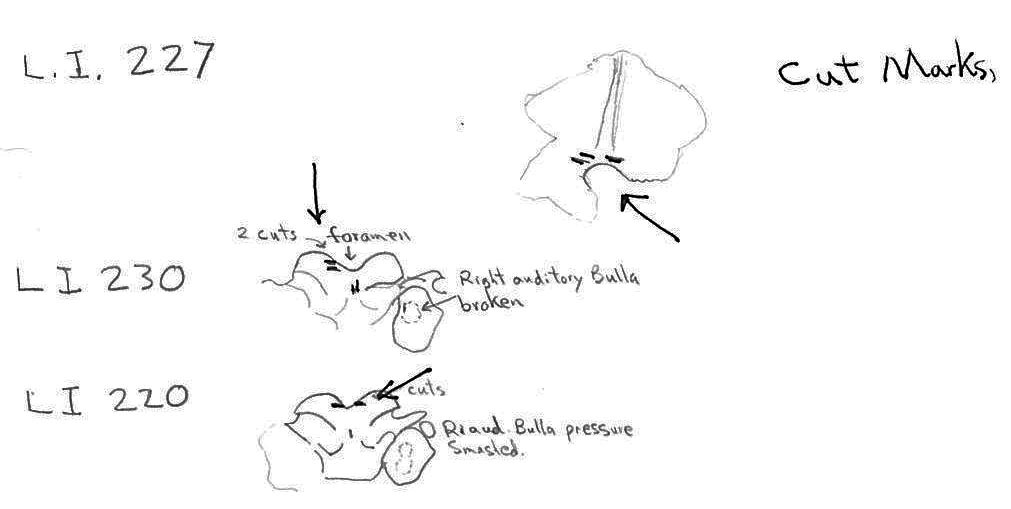
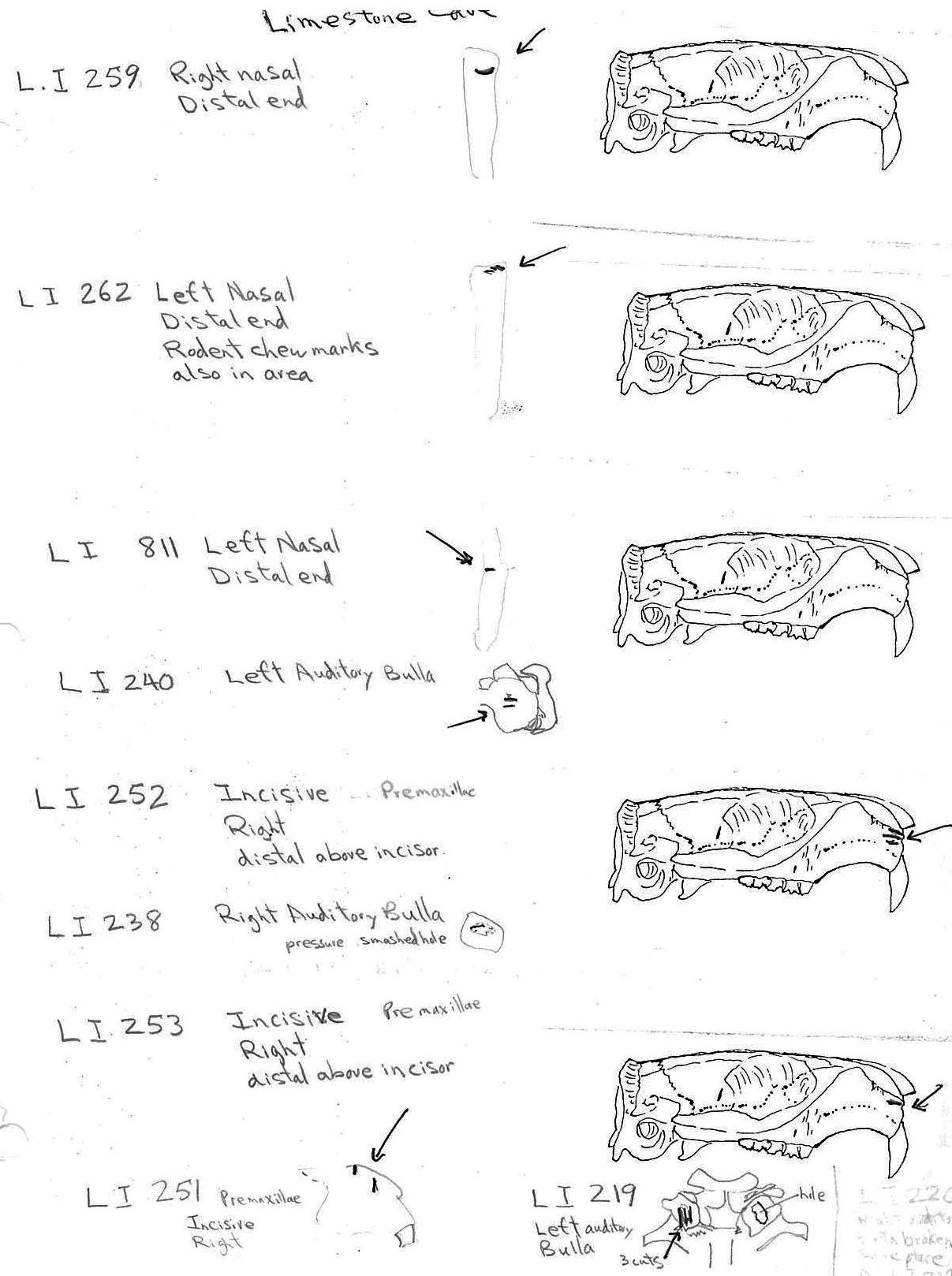
The craniums from Limestone Rock shelter are highly fragmentary. The bones at this site are semi-exposed and most are lying on or into a litter mat of fir needles. No cranium is complete. In addition to the three larger parts (LI219, LI220 and LI222) that count as individuals, there are 6 occipital bones that are from additional individuals, making the MNI cranium count nine. There are 7 left and 6 right bullae and 6 right nasals.
Of the three partial craniums that count as individuals, two have cut marks and another 10 cranium parts have cut marks. A minimum of 4 individuals are represented by the 12 specimens with cuts. The 12 modified specimens include evidence of skinning in the form of cuts on two left and one right nasal and three right incisives. The cutting of the tongue muscles is indicated by cuts on a left auditory bulla that also has a punctured right bulla, and cuts on an individual left bulla. Three other right bullae have puncture marks. Three specimens have cuts across the edge of the foramen magnum indicative of severing of the head.
Modification Of The Mandible
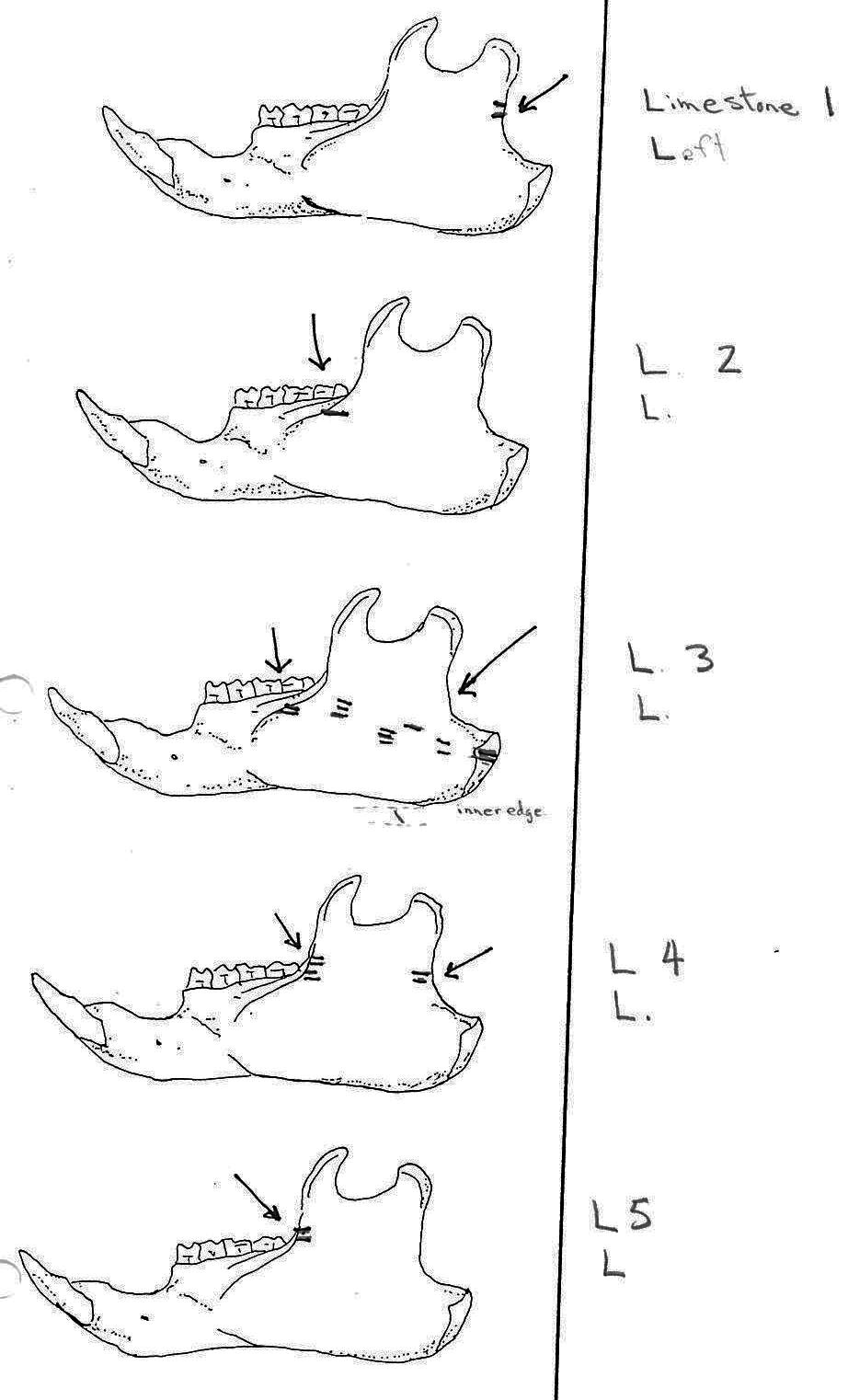
Of 74 mandibles, 27 have cuts and 16 are missing parts where cut marks usually occurred or are too corroded. Out of a visual potential of 58 mandibles only 31 are therefore complete enough to say for certain that they have no cut marks. Twenty of these mandibles without cuts are those of smaller animals. This suggests that cut marks are more common on bones when there is a need to cut the heavier muscles of the larger animals.
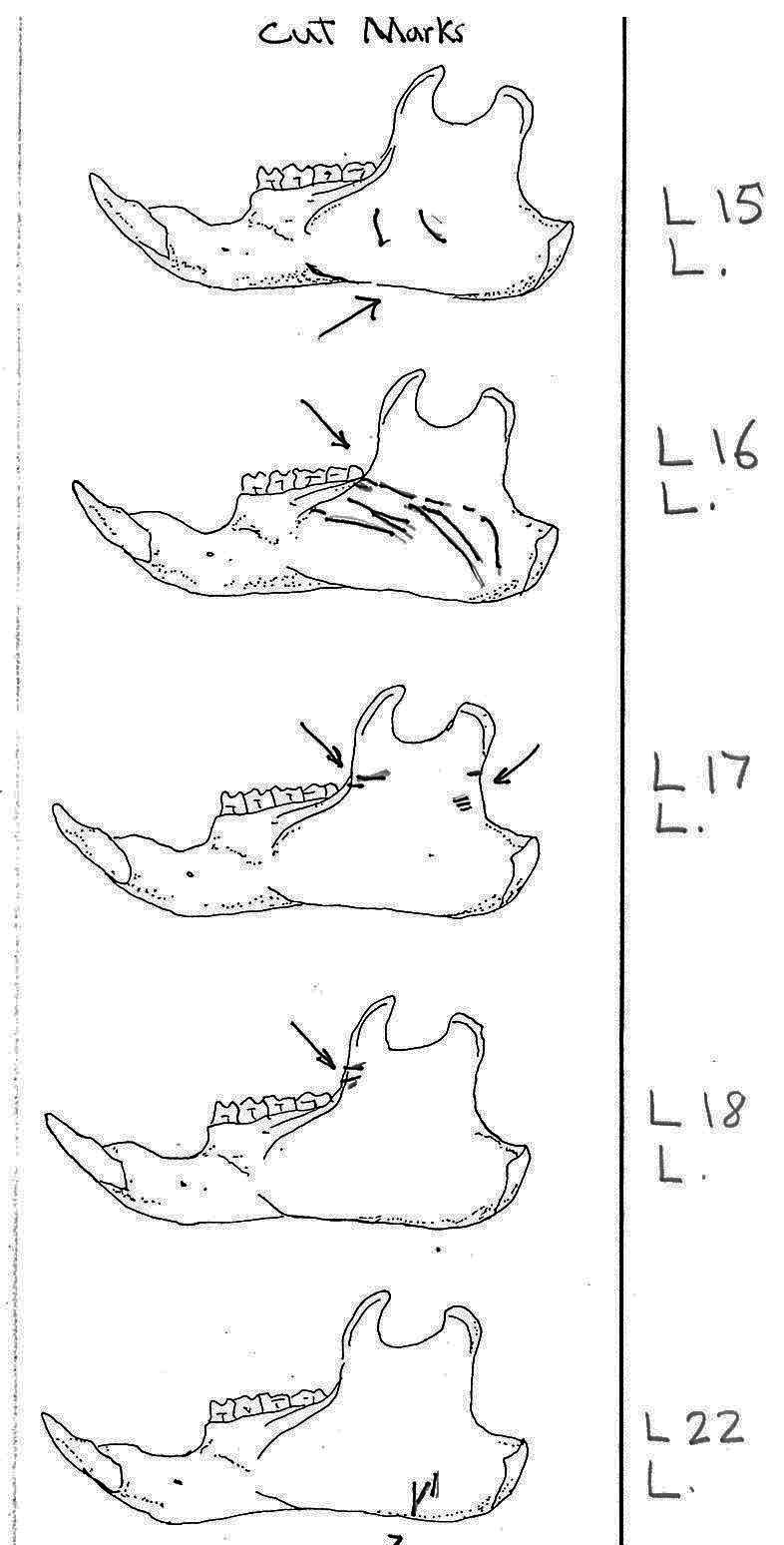
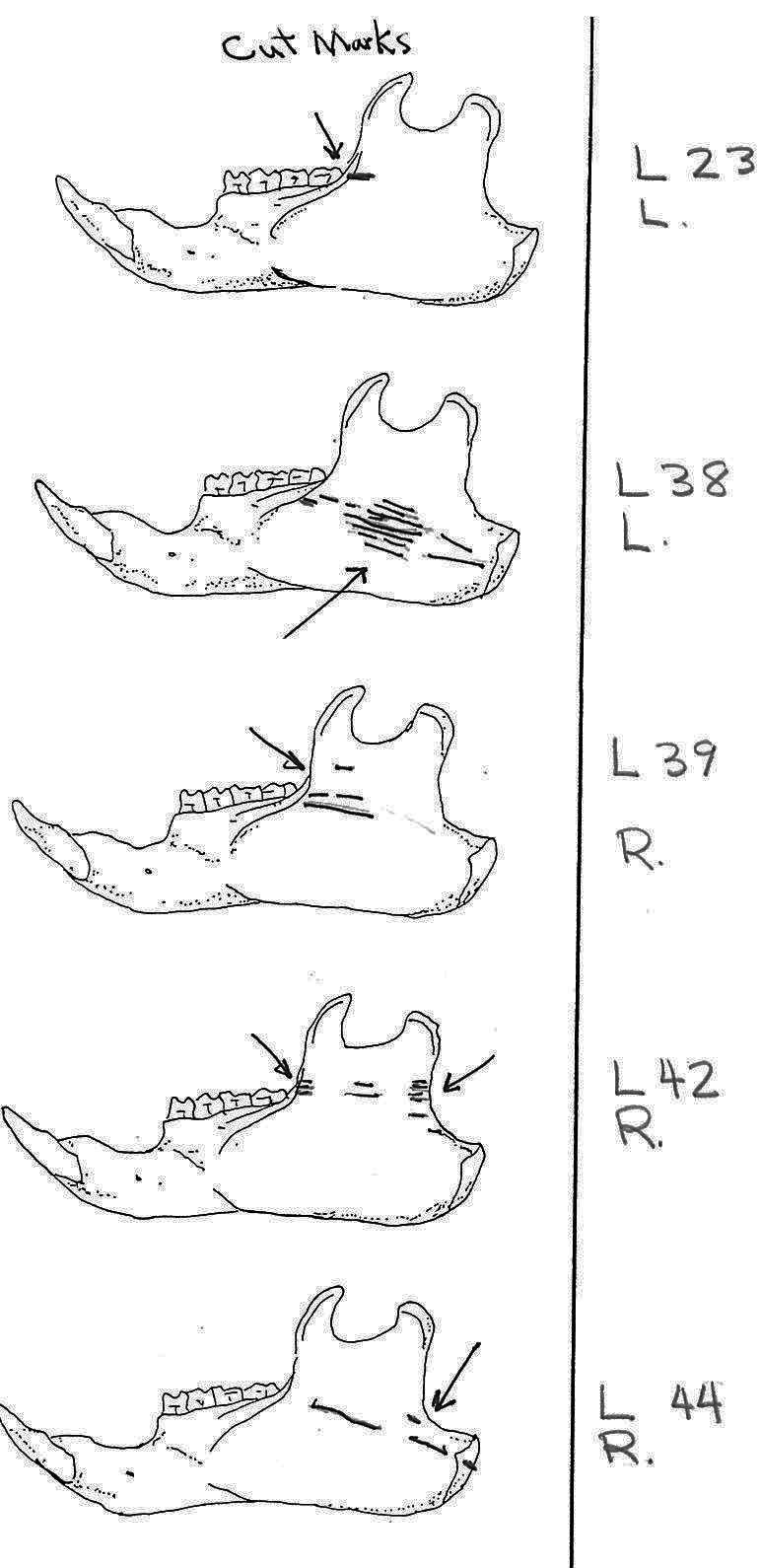
Of the 36 right mandibles 10 have butcher marks, and of the 38 left mandibles 17 have butcher marks. Most common are cuts across the ascending ramus resulting from the cutting of the masseter muscle. Fourteen have cuts on both sides of the ramus, with half of these having more cuts across the centre of the ramus. Six have cuts only near the anterior edge and two only near the posterior edge of the ramus. L3 and L11 have cuts on the underside of the posterior part of the ramus and in addition L3 has many cuts across the centre of the ramus. L7 is different in having cuts on the posterior of the main ramus as well as four cuts on the bottom of the inner or labial side of the main ramus. L15 has two curving downward cuts in the middle of the ramus and L22 has three downward cuts on the lower side of the main ramus. L54 which has the standard cuts across the ascending ramus also has cuts at the middle of the labial edge of the main ramus. L53 is different in having cuts on the labial side across the upper ramus, as well as the middle and lower body of the ramus. The cuts on the labial side of some specimens are likely secondary to the cutting involved in the removal of the tongue.
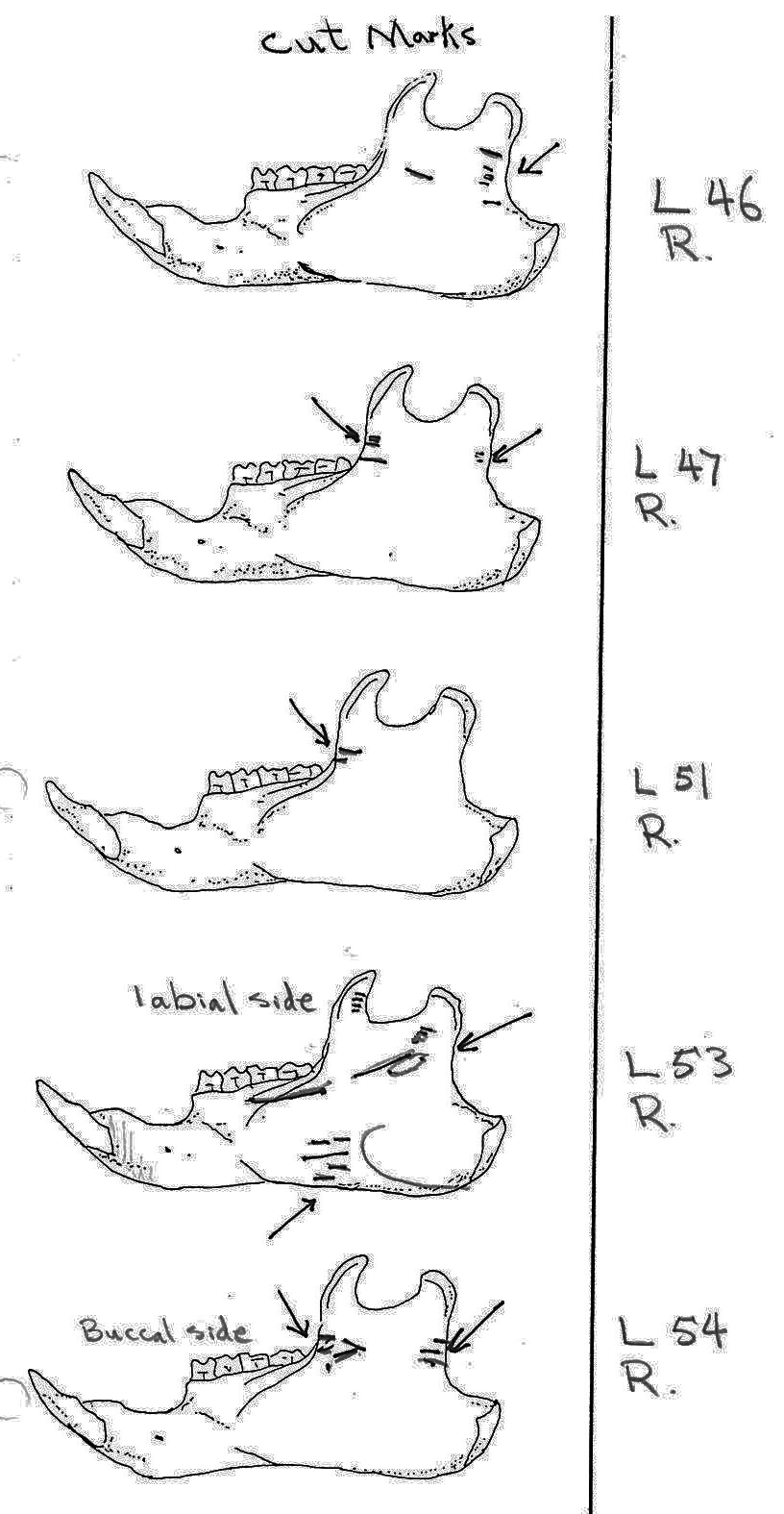
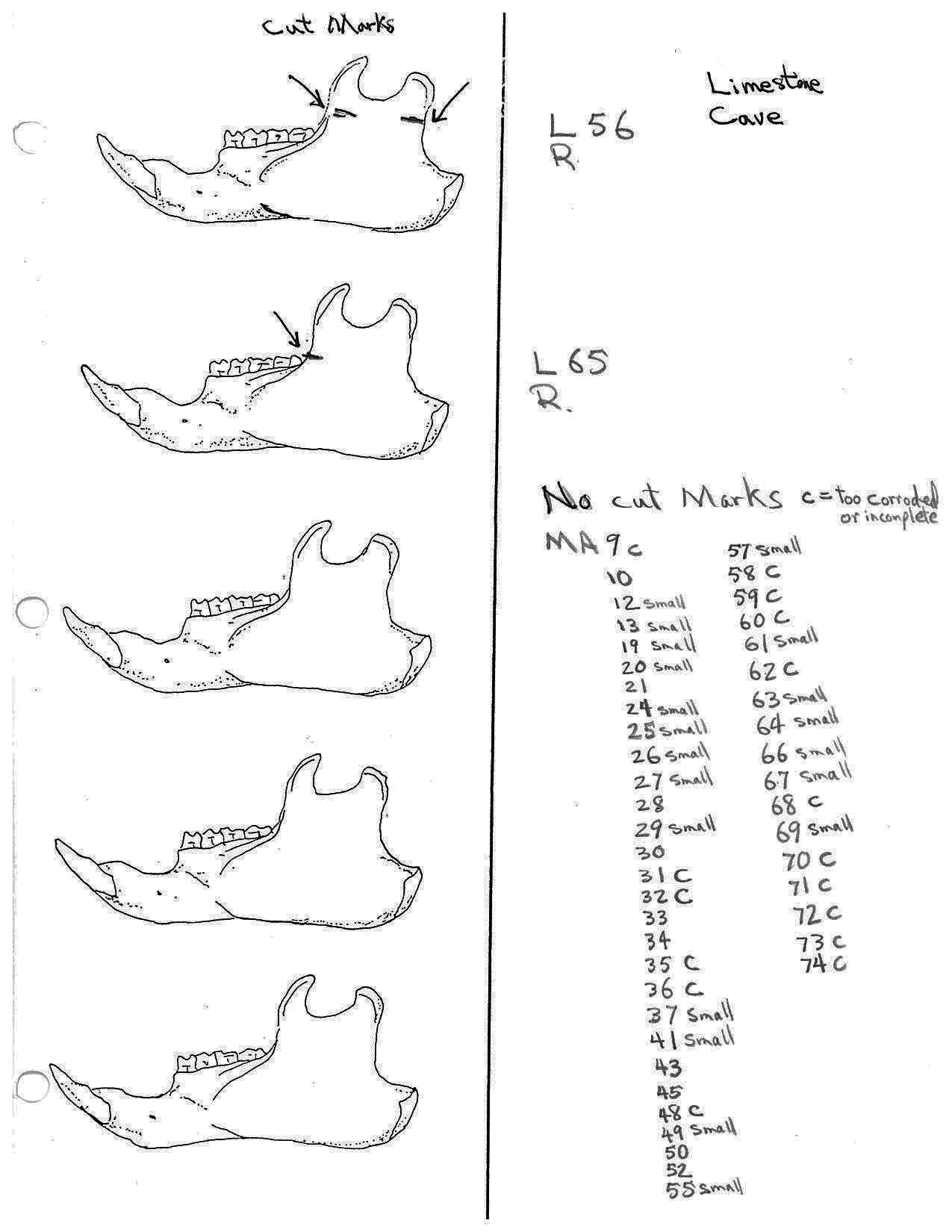
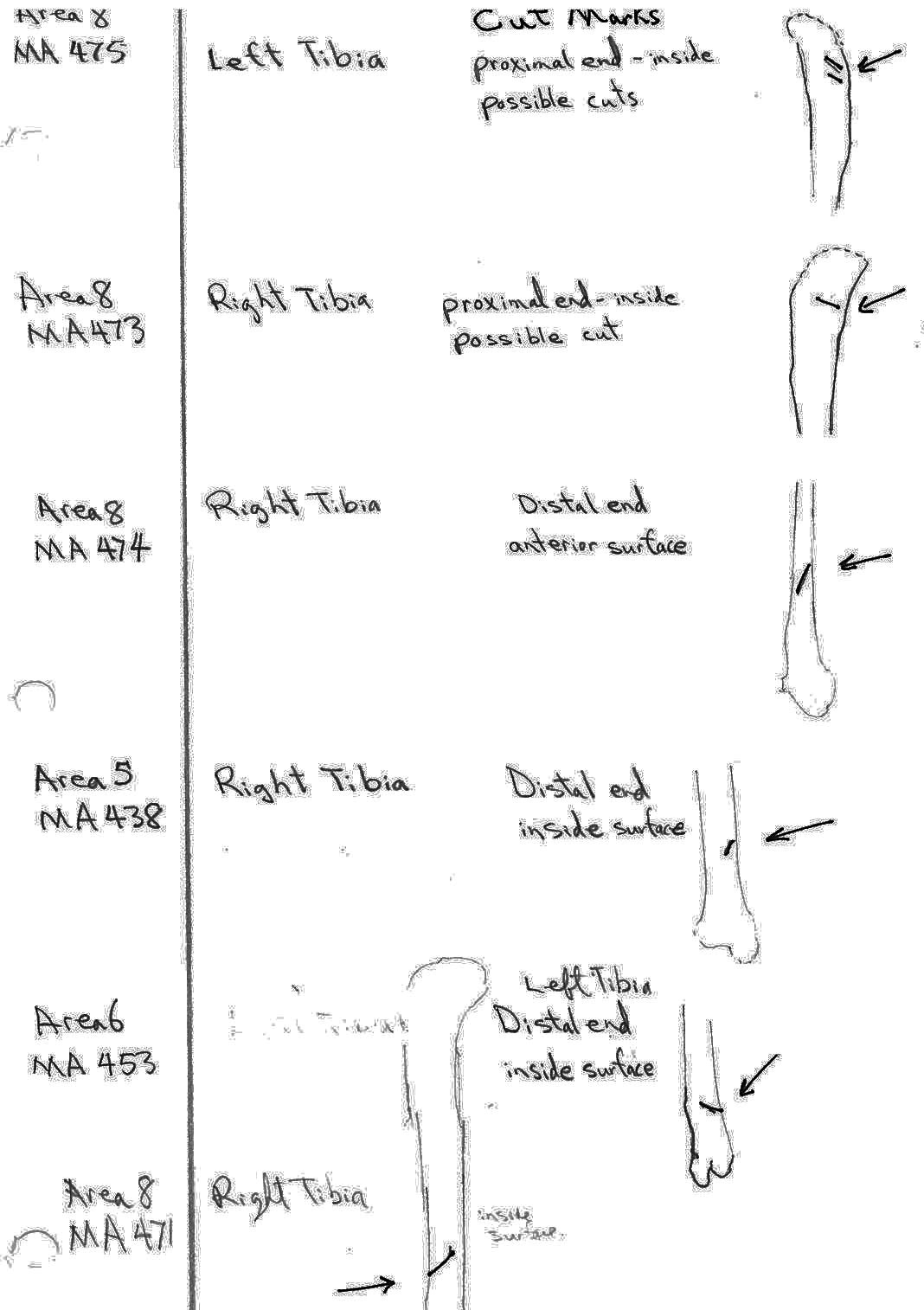
There are 7 right and 7 left femurs with cuts. All are cut near the proximal end and one is also cut just above the distal end. Eleven have the cuts only on the neck of the femur; two have cuts on the neck and across the upper greater trochanter and one is cut just below the outer trochanteric crest. The proximal cuts are consistent with the severing of the leg. The one distal cut may be related to removal of the fur from the leg area.
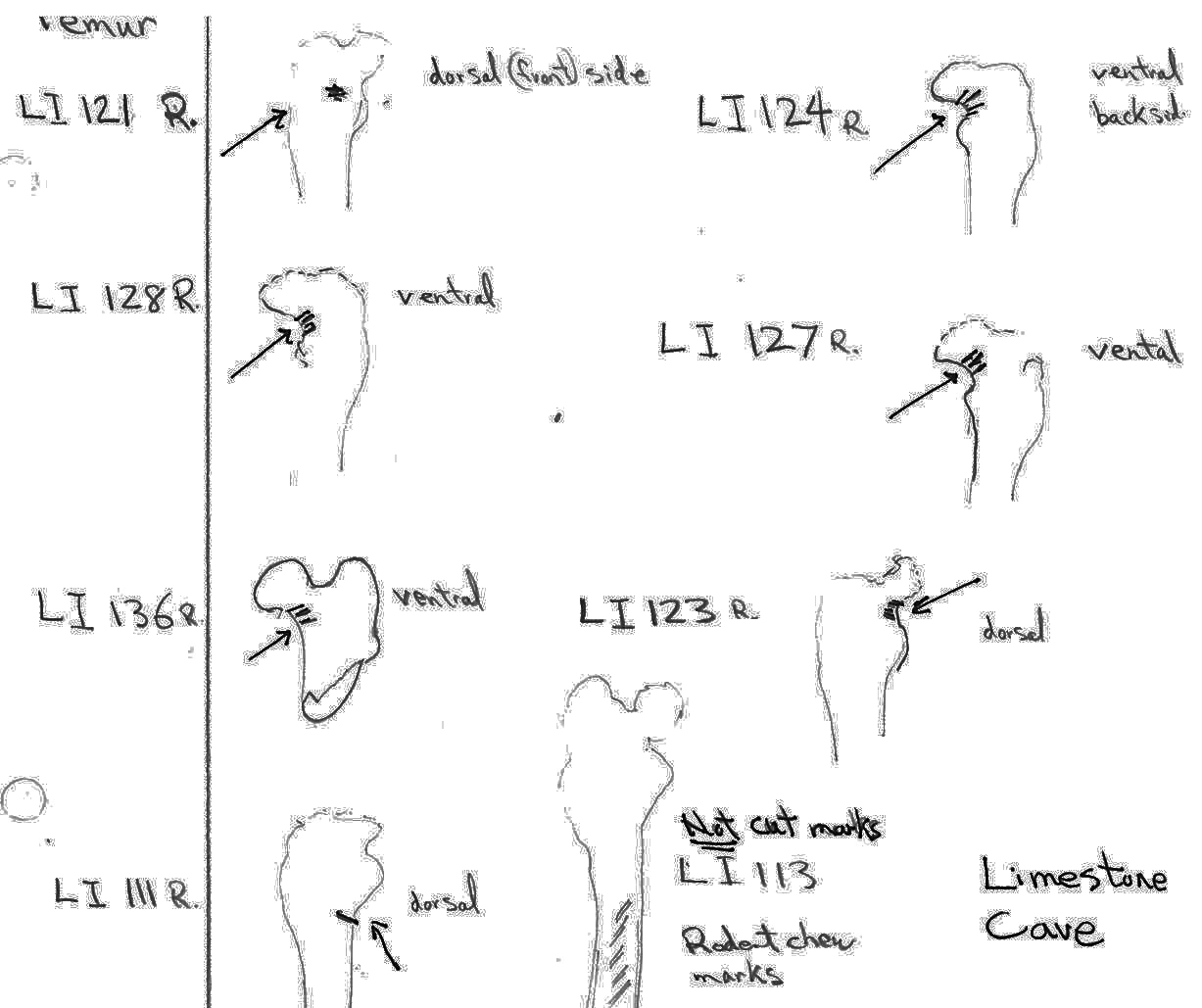
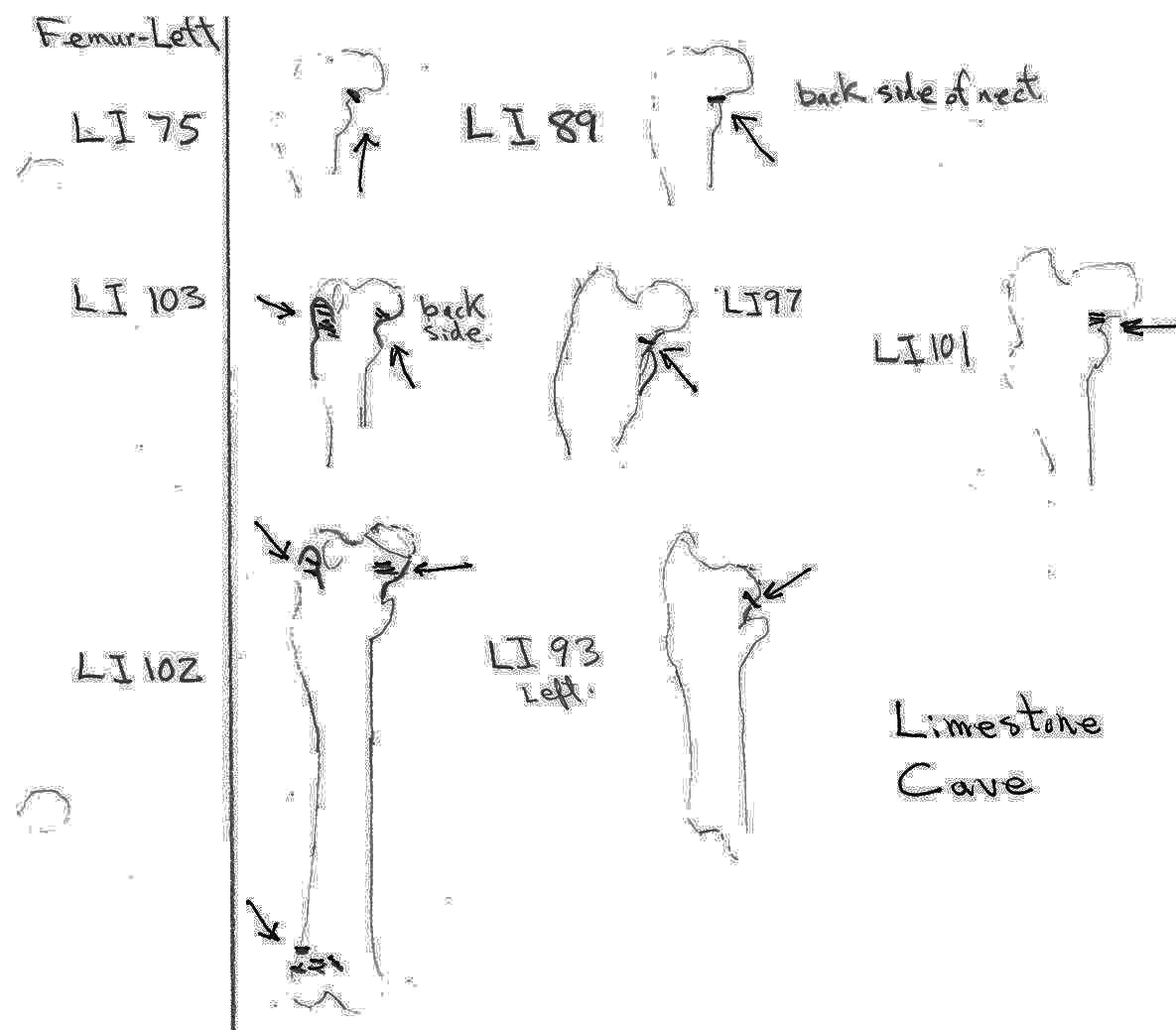
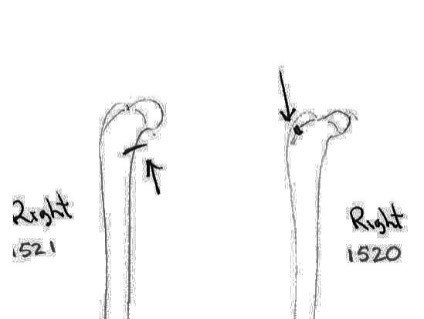
Modification Of Tibias
Only four tibias have cut marks that do not clearly represent a pattern. Two have single angled cuts near the distal end and one has cuts across the side of the distal end. One has a single long cut on the inside of the proximal shaft. These cuts seem more likely to be related to skinning than butchering.
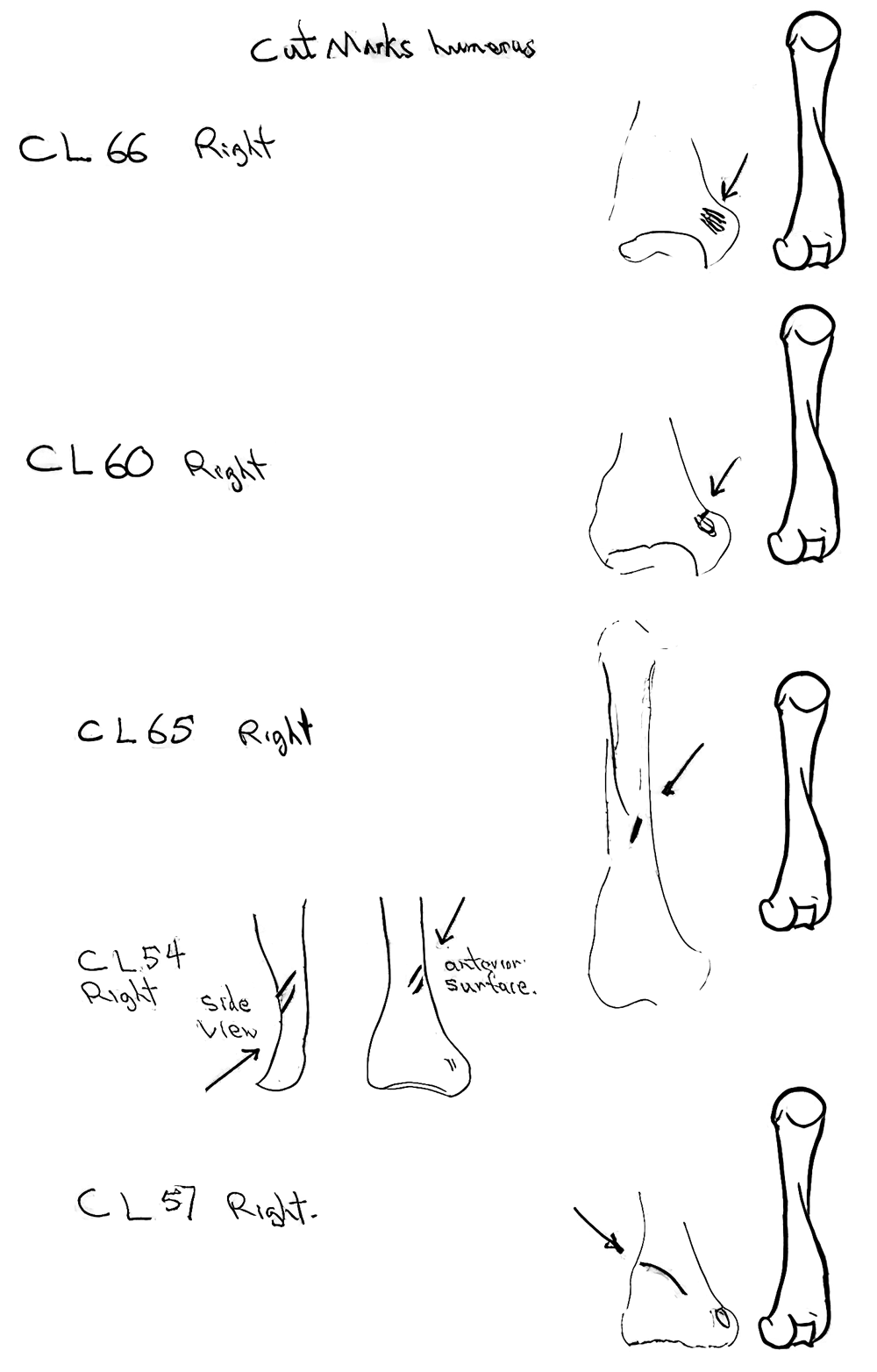
Modification Of Humerus
Twelve humeri (8 left and 4 rights) are all cut only on the distal end. Seven have cuts on one edge of the dorsal surface across or near the small bone ridge above the lateral epicondyle on the anterior surface. One has cuts in a similar location of the ventral side. These cuts are the result of cutting muscles and suggest the removal of the front paws, possible in the skinning process. Three of the humerus have single cuts angling down on the distal end of the shaft and one has cuts across the same area. These cuts seem consistent with removal of the skin from the lower forelegs.
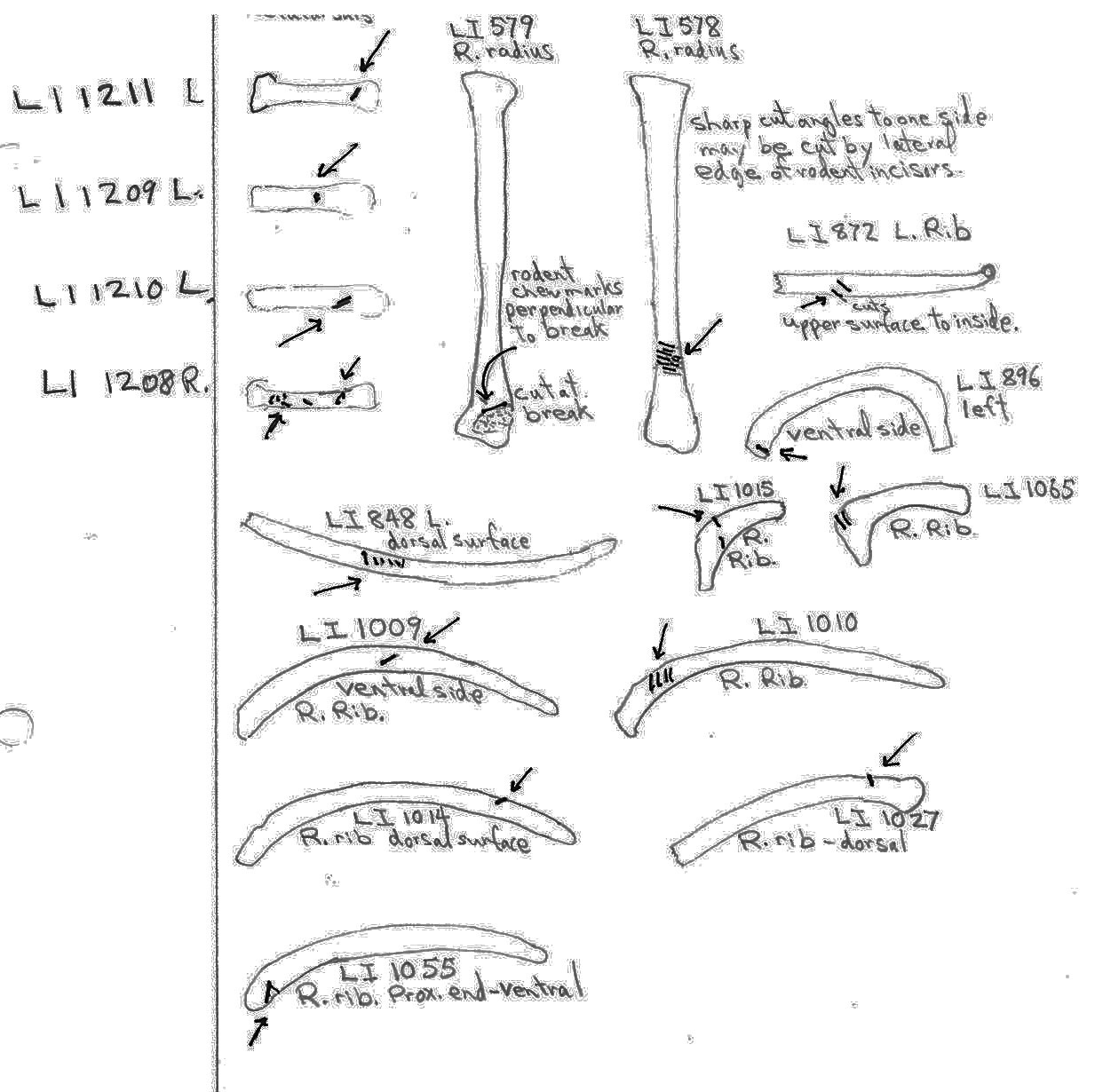
Modification Of Ribs
Ten ribs have cut marks which are atypical given the general absence of ribs from the site. Four are cut near the proximal end, three near the distal end and three near the middle. The distal end cuts may have been made during the separating of the belly fur but the others are suggestive of the filleting of the ribs.
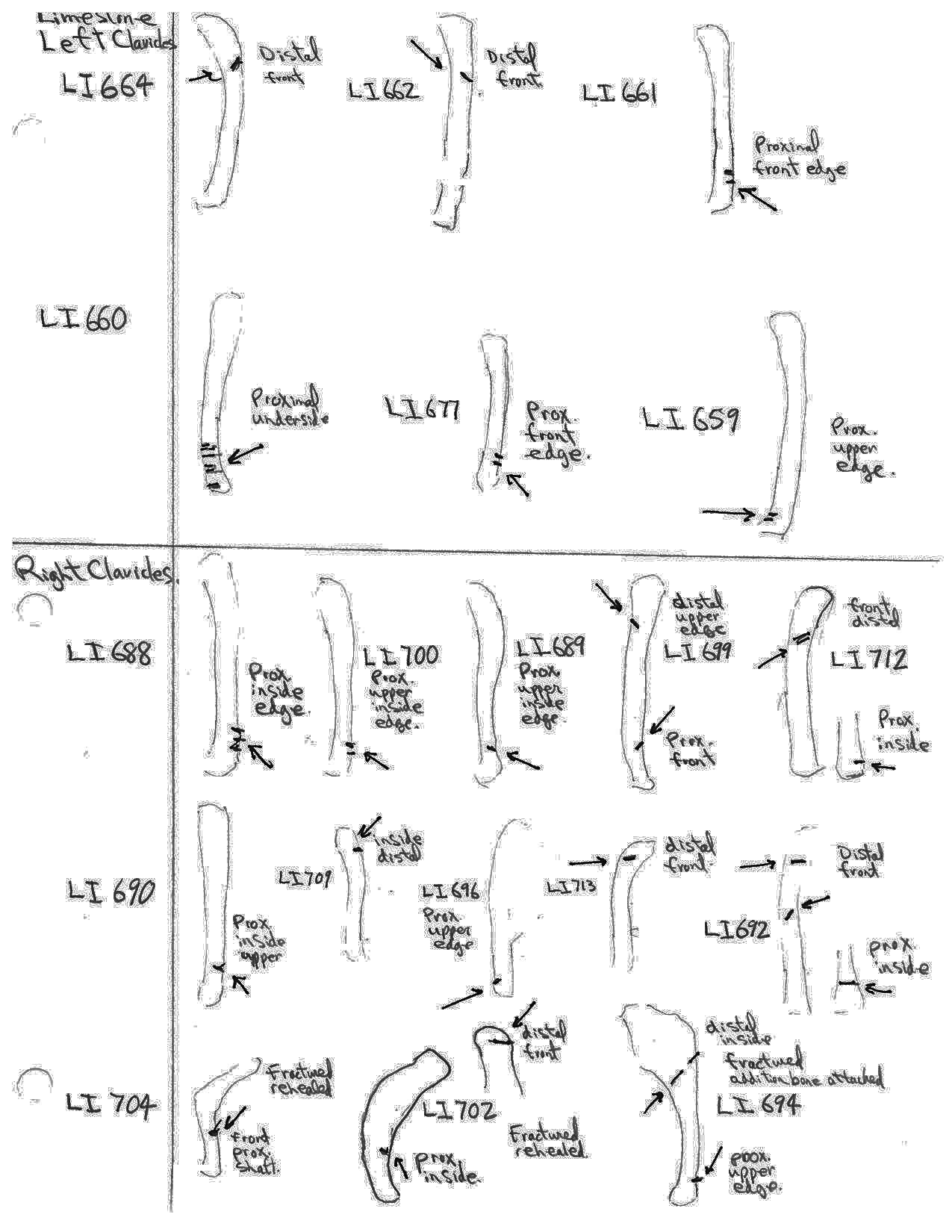
Modification Of Clavicles
Nineteen clavicles (13 right and 6 left) have cuts that are mostly toward the distal or proximal ends and occur on both anterior and posterior sides. Four have cuts near both ends. These cuts are consistent with the removal of the front limbs. The scapula (which have no cut marks) may have been removed at this time.
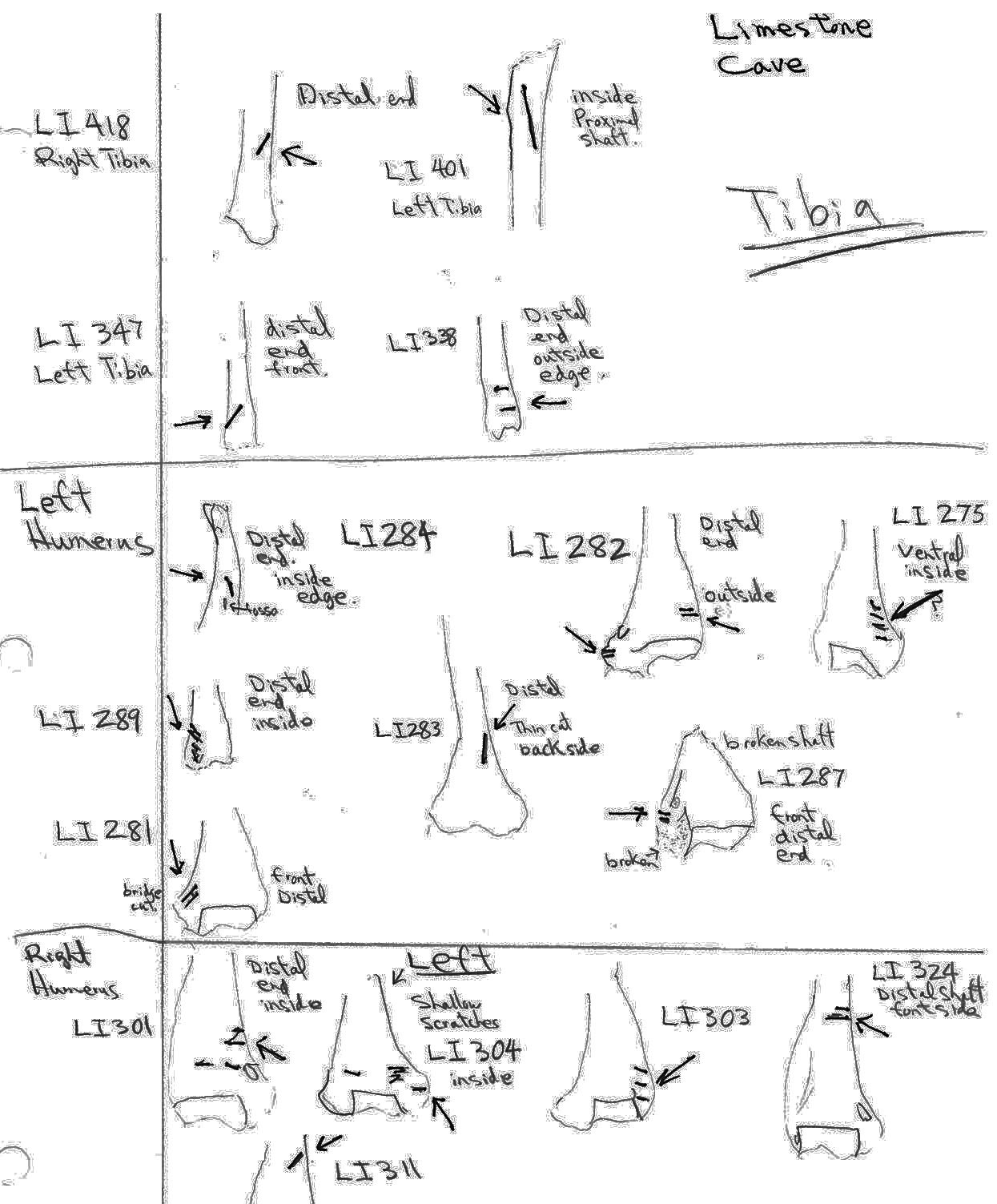
Modification Of Radius
Only two right radiuses have distal end cut marks and both are not clearly defined. One has a series of cuts with sharp angles to one side which could potentially be caused by the lateral edge of a rodent incisor. The other appears to be an angled cut on a broken edge, which also includes rodent chew marks perpendicular to the break. These are inconclusive to establish a pattern but may be related to skinning during removal of the forearms.
Modification Of The Metatarsal
Three left metatarsal have single cuts near the distal end and one right specimen has several random cuts on the shaft. These would be related to skinning and not butchering.
Marmot Bones From Limestone Cave
SUMMARY LIST OF PRESENTS AND ABSENCE OF CUT MARKS
(MNI = Minimum number of individuals)
(Showing MNI represented by each cut element class and the total MNI in the site represented by that element)
| BONE ELEMENT: | NUMBER WITH CUT MARK(S) |
|---|---|
| Mandible | 27 (MNI CUT 17) (site MNI 74) |
| Cranium (and parts) | 12 (MNI CUT 4) (site MNI 9) |
| Femur | 14 (MNI CUT 7) (site MNI 7) |
| Tibia | 4 (MNI CUT 3) (site MNI 52) |
| Humerus | 12 (MNI CUT 8) (site MNI 36) |
| Ribs | 10 (MNI CUT 1) (of 376 ribs) |
| Clavicle | 19 (MNI CUT 13) (site MNI 13) |
| Metatarsal | 4 (MIN CUT 1) (site MNI 1) |
| Radius | 2 (MNI CUT 2) (TOTAL MNI 51) |
BONE ELEMENTS WITH NO CUT MARKS: Fibula, scapula, ulna, innominate, vertebrae.
Conclusions Deduced From The Absents Of Bones And Modifications
The marmots were skinned around the nose and lower legs. The tongue was cut out, the masseter muscle cut and the head pulled back and severed at the base of the cranium from the front.
The back legs were cut off from the pelvis, possibly to facilitate the removal of the pelt. The absence of cut marks on the pelvis suggests that the softer tissue may have been cut and the abdominal area snapped back and pulled off. In some cases the front lower legs were cut, also to facilitate skinning, and the ulna and radius pulled out after separation of the lower leg, leaving the foot bones attached to the pelt.
The upper humerus has no cut marks and may have been pulled off with the scapula after the cutting of the shoulder muscles down to the clavicle. The entire back bone and rib cage were taken away from the site with the exception of those of a few animals whose rib cages seem to be present at the site.
Modifications To Non-marmot Bones From Limestone Cave
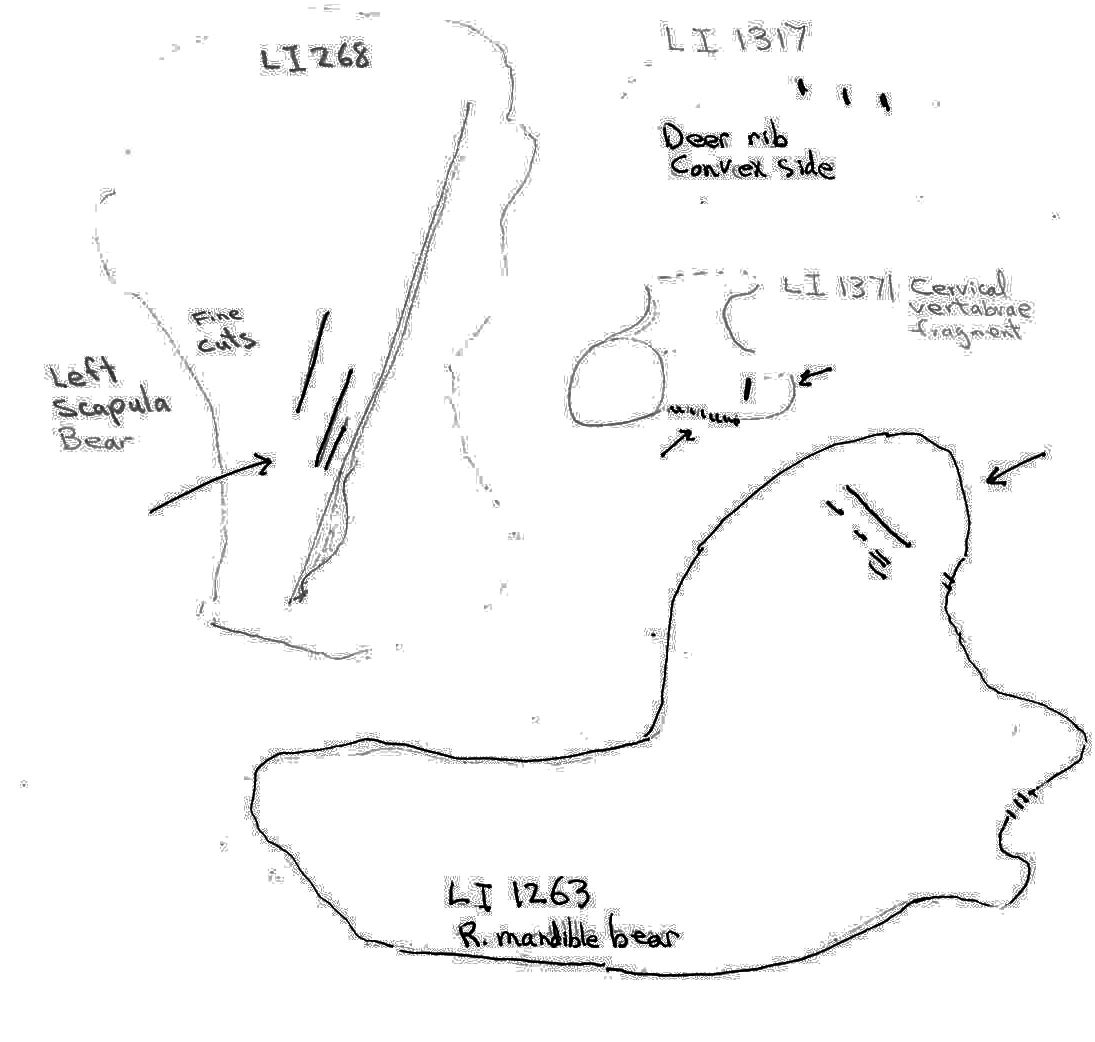
Non-marmot bones from Limestone Cave were highly fragmentary. Cut marks were observed on two bear and two deer bones. On a bear right mandible (LI1263) 3 cuts occur on the lower base of the posterior of the condyloid process and several cuts occur on the posterior and midsurface of the coronoid process of the ascending ramus. Three long, fine cuts were found on the dorsal surface of a left bear scapula (LI268) parallel to the spinous process.
A deer rib mid-section fragment (LI1317) has 3 cuts on its dorsal surface and a deer cervical vertebra fragment (LI1371) has 9 short close together cuts and one near by cut on the edge of one side.
There are not enough cut bones to note any standard carcass processing techniques. The fragmentary nature, incompleteness, and few examples of butchered remains that do occur indicate that deer and bear were heavily butchered and parts removed from the cave site.
The Golden Hinde Rockshelter Bones
Only marmot bones occur at this site. The remains represent a minimum count of 4 individuals. Most bones were examined in the field and left at the site. Initially Lindsay Elms of Campbell River collected a left mandible with cut marks on each side of the base of the ascending ramus. He also collected a R. scapula; R. Maxillae fragment; L. and R. incisors and one L. and two R. incisive fragments.


Figure 8. Entrance to Golden Hinde Rockshelter. Left to right Linday Elms, Mark Dickson and Grant Keddie. Dave Nagorsen photograph October 4, 1994.
Marmot bones were found in two distinct areas 1 and 2 along the east wall of the rock shelter. Two separate bones formed area 3 on the opposite side of the compartment floor. Four small bones lay on the top of a rock near the entrance to the cave. These appear to have been picked up from the cave floor and placed here in recent times, probably by hikers examining the bones. It is likely that some of the larger bones, especially skulls, may have been removed by recent visitors.
The bones were in a heavily weathered and fragmentary condition making it difficult to identify cut marks. Three left and three right mandibles indicate there were at least three young of the year marmots. A larger tibia with a partially fused epiphysis indicates the presence of an additional juvenile.


The field observations revealed only one more cut bone in Area 2. This is a rib with missing ends that exhibits three cut marks – one across the rib near the proximal end and two adjacent cuts across part of the rib. The sparse evidence from this site indicates that at least some butchering occurred here, but the presence of a vertebral column and ribs in area 2 may suggest at least one animal was skinned and no parts taken away for food consumption.
Bones in Area 1 included: L. maxillae fragment with tooth sockets; 2 L. ribs; L. radius; L. ulna; L. fibula; L. scapula; L. humerus fragment; L. mandible with two molars; 5 small unidentified long bone fragments; R. humerus; R. ulna; R. innominate with missing pubis; L. Clavicle; 2 R. tibia (different sizes); L. radius; R. radius fragment; R. mandible fragment; 2 L. tibia fragments.
Bones in Area 2 included: 2L. and 3R. mandibles (with incisors); a L. and R. incisor; 15 ribs (side not identified); 2 fragmentary fibulas (side not identified); R. radius; L. femur; L. tibia (distal end partly fused); epiphysis for left tibia; R. humerus; 3 R. pubis; L. pubis; 3 R. and 3 L. ilium; L. and R. matching maxillae fragments with tooth sockets; L. and R. matching frontal fragments; parietal fragment; 12 thoracic vertebra and 5 lumbar vertebra;
Bones in Area 3 included: L. innominate (missing part of ischium and pubis) and a small rib fragment.
Radiocarbon Age Estimates On Marmot Bones
Most samples included here are Isotrace Lab ATM dates on individual femurs or a tibia in the case of Clayoquot (TO-692) and a mandible from the Golden Hinde. Also included are standard radiocarbon estimates from the Shoemaker Bay site on charcoal found in layers containing marmot bones.
SITE DATES. ISOTRACE LAB #. AGE (Years BP). CALIBRATED
| CLAYQUOT | TO-1224 | 2630+-50 | B.C. 807 |
| TO-692 | 2490+-50 | B.C. 600 – 680 | |
| MARINER | TO-3562 | 890+-50 | 1163 A.D. |
| TO-3563 | 920+-50 | 1047-1154 A.D | |
| TO-3564 | 990+-50 | 1022 A.D. | |
| TO-3565 | 850+-50 | 1211 A.D. | |
| TO-3566 | 890+-50 | 1163 A.D. | |
| TO-3567 | 970+-90 | 1027 A.D. | |
| LIMESTONE | TO-4266 | 1030+-60 | 1015 A.D. |
| TO-4267 | 1070+-60 | 990 A.D. | |
| GOLDEN HINDE | TO-4265 | 830+-60 | 1225 A.D. |
| SHOEMAKER BAY | – | 1450+-80 | 609 A.D. |
| – | 1130+-85 | 894 A.D. |
The two dates from Clayoquot span a time range of 207 years. The two samples were dated at different times. This suggests that some unknown laboratory procedure may have been a factor in explaining the difference in the dates obtained.
The six dates from the Mariner site span 189 years and the two dates from Limestone Rockshelter span 25 years. When the Golden Hinde site is included with the latter two sites there is only a time range of 235 years for all three sites. The layers of the Shoemaker Bay site containing marmot bones span a time range of about 285 years.
Index 1
Summary Of Marmot Bones From The Shoemaker Bay Site
The Shoemaker Bay site, recorded as DhSe- 2, is an aboriginal shellmidden located on Alberni Inlet west of Port Alberni. A total of 42 two meter square units and 3 small units were excavated at this site – 27 in 1973 and 18 in 1974. Separate studies of the faunal remains were undertaken for materials collected in each of the years. The site is composed of two major components or zones. All of the 24 marmot bone elements recovered can be considered as being found in the upper Component II or Zone A, although 4 are from an uncertain zone A/B. Zone A (Levels 1-4) is radiocarbon dated to 1450+-80 B.P. (Gak5108) [corrected date 1341 B.P. or A.D. 609] and 1130+-85 B.P. (Gak5432) [corrected 1056 B.P or A.D. 894].
Information on the type of bone elements for the 1973 faunal material is missing due to the loss of the original material. Before they were lost, I documented the number of elements and provenience of each. Material was collected in 10cm. level increments. If the minimum number count is judged by each level, there are a minimum of 18 marmots represented. Seven of the 45 excavation units contained marmot bones in 12 different levels.
The 1974 Faunal Material
Sources: Report on DhSe-2 Faunal Analysis (Port Alberni Site) University of Toronto student manuscript, by Leigh Field and Bernadette Laqueur, May 23, 1975.
This study included the analysis of mammalian bones recovered from 18, two meter square, units in 1974. Out of 3625 mammal remains 912 or 25.2% were identified as to species. Most specimens were identified from the upper three levels (1 to 3) with a total of 768 bones. This compares with 144 bones identified for the next five lower levels (4 to 8). Marmot remains are found in 3 of the 18 units and in six levels.
A total of 10 marmot elements were found representing 1.1% of the identified mammal elements. Six elements were found in levels 1-3 (.8% of mammals) and 4 (2.8%) in levels 4-7 [all from level 4].
Marmot remains by level with total for mammal remains in brackets: level 1 – 0 (266); level 2 – 2 (322); level 3 – 4 (182); level 4 – 4 (67); levels 5 to 7 – 0 (75). Marmots are not found in the bottom 3 levels and starting in level 4 represent 6%, 2.2%, .6% and 0% as the levels move up through time. Although these statistics show a reduction in marmot elements through time the size of the sample is too small to assume any significance to this trend. The information available shows a minimum of 8 individuals when counting the minimum number per level.
The marmot elements identified include the following – given unit number, level number and bone number: I2-12, L. humerus; I4-1, L. humerus; J3-23, R. innominate; J3-24, R. innominate; J3-25, L. scapula; J4-1, R. humerus; J4-7, R. humerus; J4-8, R. temporal; N2-1, L. humerus (unfused epi.); N3-1, L. humerus (proximal epiphisis unfused and cut marks medial to lesser trochanter).
1973 Excavation Results
Source: Alberni Prehistory. Archaeological and Ethnographic Investigations on Western Vancouver Island, by Alan D. McMillan & Denis E. St. Claire, Theytsus Books, Penticton, 1982.
Excavation of twenty nine 2m square units produced the following number of bones: component I (1458 bones); I/II (4277); III (10629). This included the recovery of 4389 mammal bones (31% by bone count). Family identification by component is: I – 17%; II – 35%; I/II – 30%. Mammal species by component included Odocoileus hemionus I 85%, II 82 %; Ursus americanus I 5%, II 1%; Martes americanus II .5%.
Component I
No marmot bones are from this component. Of the 654 mammal bones 112 or 17% were identified to species. Deer bones (75) comprised 67% of the sample, a minimum number of 6, or 32% of the total minimum numbers; 2 black bear are 10.5% of total minimum number.
Component II or Zone A and part of Zone A/B
When counting the minimum number of individuals per level this represents 8 individuals. This is the same minimum number for the similar 1974 sample of 1066 identified mammal bones. Deer (537 bones) include a minimum of 25 individuals or 43% of total of 59 MNI. Marmot remains included 14 bone elements from 6 levels of 4 units – 10 bones from Zone A (MNI 5) and 4 (MNI 3) from Zone A/B. These are from the following Zone, unit and level:
Zone A – E14-16 S6-8, Level 2, A/1.90 – A/2.00 – 7 elements (2 individuals). [All other levels represent one individual].
Zone A – E18-20 S2-4, Level 2, A/1.75-A/1.85 – 1 element.
Zone A – E18-20 S6-8, Level 3, A/1.90-A/2.00 – 1 element.
Zone A – E12-14 S8-10, Level 1, Surface to A/2.00 – 1 element.
Zone A/B – E12-14 S8-10, Level 2, A/2.00 – A/2.10 – 2 elements.
Zone A/B – E12-14 S8-10. Level 3, A/2.10 – A/2.20 – 1 element.
Zone A/B – E14-16 S6-8, Level 3, A/2.00 – 2.10 – 1 element.
Index 2. Cut Marks On Modern Laboratory Marmot Specimens
For comparative purposes I examined six marmot skull specimens in the existing Royal B.C. Collection that were processed in the field or lab using steel scalpels or knives. Only some of these were in good condition for preserving the pelt when they were processed. Specimen 018546 was in good condition when it was skinned. It exhibits skinning cut marks across the ascending ramus of a left mandible and an incisive cut below the nose area similar to processing marks on the ancient specimens from Mariner Cave and Limestone Rockshelter.
Other collection specimens do not exhibit similar cut marks to the ancient specimens. These can not be used as comparative examples because of the differing condition in which the animals were found. Some of the specimens already partially skeletized. Some specimens show more extensive cutting on the rostrum and incisives which is probably due to removing tighter and dryer skin. Specimen 5021 has many cuts across the rostrum and on both incisives. Specimen 10709 has several cuts across the rostrum and several above the rostrum and one on the left incisive. Specimen 1260 has a series of cuts below and perpendicular to the tooth row on the right mandible which is likely a result of removing tightly adhering skin. The base of this skull is cut in one direction on the occipital condoyle and in the opposite direction across the basio-occipital. Specimen 11179 has four cuts similar to 1260 on the occipital condoyle. Specimen 15307 does not exhibit cuts.